Evidence for a novel feedback loop in the Hedgehog. pathway involving the seven transmembranedomain protein Smoothened and the kinase Fused..
|
|
|
- Marvin Bell
- 6 years ago
- Views:
Transcription
.")


1 Evidence for a novel feedback loop in the Hedgehog pathway involving the seven transmembranedomain protein Smoothened and the kinase Fused. Sandra Claret, Matthieu Sanial, Anne Plessis To cite this version: Sandra Claret, Matthieu Sanial, Anne Plessis. Evidence for a novel feedback loop in the Hedgehog pathway involving the seven transmembranedomain protein Smoothened and the kinase Fused.. Current Biology - CB, Elsevier, 2007, Sous presse (?). <hal > HAL Id: hal Submitted on 13 Jul 2007 HAL is a multi-disciplinary open access archive for the deposit and dissemination of scientific research documents, whether they are published or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
2 * Manuscript This manuscript has been accepted for publication in Current Biology Evidence for a novel feedback loop in the Hedgehog pathway involving the seven transmembrane-domain protein Smoothened and the kinase Fused. Sandra Claret 1, 2, Matthieu Sanial 1 and Anne Plessis 1 *. 1 Laboratoire «Génétique du Développement et Evolution», Institut Jacques Monod - Université Paris 7, UMR 7592, CNRS/Universités Paris 6 and 7 2 Place Jussieu Paris, France 2 Current address: Laboratoire «Morphogenèse et polarité», Institut Jacques Monod - Université Paris 7, UMR 7592, CNRS/Universités Paris 6 and 7 2 Place Jussieu Paris, France * To whom correspondence should be addressed. plessis@ijm.jussieu.fr Phone: Fax: Short title: Fused-Smoothened regulation. Key words: Hedgehog, Smoothened, Fused, signalling, traffic, imaginal disc, feedback loop, Drosophila. 1
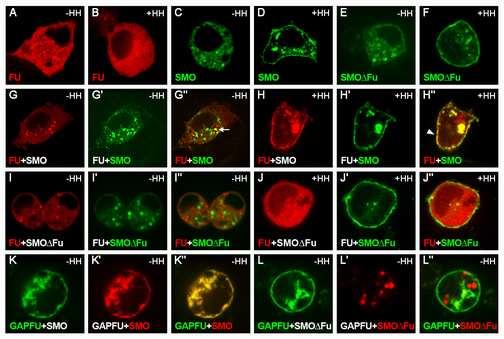
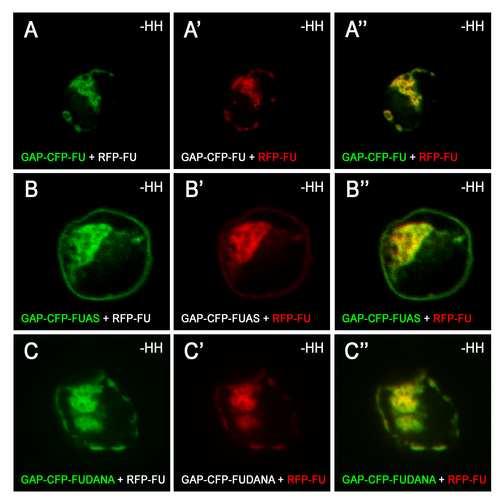
3 SUMMARY Hedgehog (HH) is a major secreted morphogen involved in development, stem cell maintenance and oncogenesis [1, 2]. In Drosophila wing imaginal discs, HH produced in the posterior compartment diffuses into the anterior compartment to control target gene transcription via the transcription factor Cubitus interruptus (CI). The first steps in reception and transduction of the HH signal are mediated by its receptor Patched (PTC) [3] and the seven-transmembrane domain protein Smoothened (SMO) [4, 5]. PTC and HH control SMO by regulating its stability, trafficking and phosphorylation (for review see [6]). SMO interacts directly with the Ser-Thr protein kinase Fused (FU) and the kinesin-related protein Costal2 (COS2), which interact with each other and with CI in an intracellular Hedgehog transducing complex [7-9]. We show here that HH induces FU targeting to the plasma membrane in a SMO-dependent fashion and that, reciprocally, FU controls SMO stability and phosphorylation. FU anchorage to the membrane is sufficient to make it a potent SMO-dependent, PTC-resistant, activator of the pathway. These findings reveal a novel positive-feedback loop in HH transduction and are consistent with a model in which FU and SMO, by mutually enhancing each other's activities, sustain high levels of signaling and render the pathway robust to PTC level fluctuations. RESULTS SMO promotes the relocalization of FU from vesicles to the plasma membrane in response to HH We produced FU tagged with RFP (RFP-FU) and SMO tagged with GFP (SMO-GFP) alone or together in Clone 8 cells (Cl-8) Drosophila wing imaginal disc cultured cells responsive to HH. In these conditions, pairs of proteins were overproduced, so other components were probably present at much lower levels. In the absence of HH, SMO-GFP 2
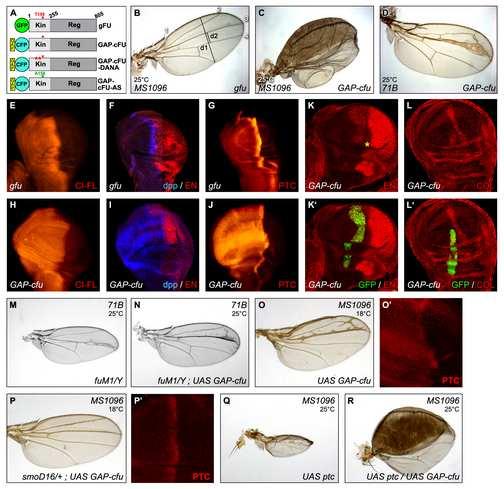
4 colocalized with RFP-FU (but not with RFP-COS2) in vesicles (Figure 1G-G, see also [9]). HH induced the accumulation of SMO-GFP at the cell surface, but had little effect on RFP- FU alone (Figure 1B, D). When both proteins were overproduced in the presence of HH, most of the RFP-FU colocalized with SMO-GFP at the plasma membrane (Figure 1H-H ). COS2 has been reported to bind the N-terminal part of the cytoplasmic tail of SMO directly, but SMO-GFP colocalized with RFP-COS2 only when HH was present (see the Supplementary Data Figure S3 of [9]). SMO FU -GFP is a mutant of SMO-GFP unable to interact with FU or to colocalize with RFP-FU in the absence of HH, due to deletion of the last 59 amino acids of SMO, which include the SMO/FU interaction domain [9]. In the presence of HH, SMO FU -GFP accumulated at the plasma membrane (Figure 1E-F) and colocalized with RFP-COS2 in the presence of HH (Supplementary Data Figure S3 of [9]), but did not recruit RFP-FU (Figure 1J-J ). Thus, upon HH stimulation, SMO-GFP induces the relocalization of RFP-FU to the plasma membrane. This effect is directly dependent on the region of SMO-GFP that binds FU and is unlikely to involve COS2. The production of a form of FU tethered at the plasma membrane disrupts normal Drosophila development and constitutively activates the HH pathway In imaginal wing discs, FU is required to upregulate genes responsive to high levels of HH [10-12]. We investigated the consequences of FU relocalization to the membrane, by producing a fusion between the N-terminal end of CFP-FU and GAP43 (GAP-CFP-FU) - a palmitoylated domain thought to anchor proteins in raft domains [13, 14] - in wing imaginal discs. As wild-type FU [15], GFP-FU accumulated throughout the wing imaginal disc (Supplemental Data Figure S1) and its overproduction of had no visible effect on wing 3
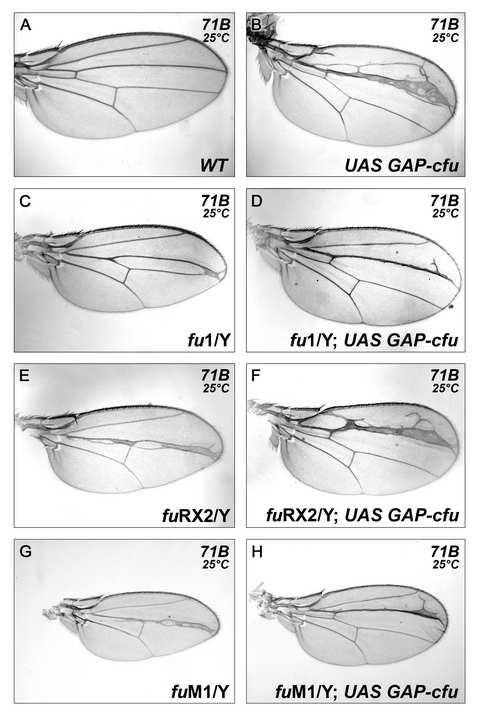
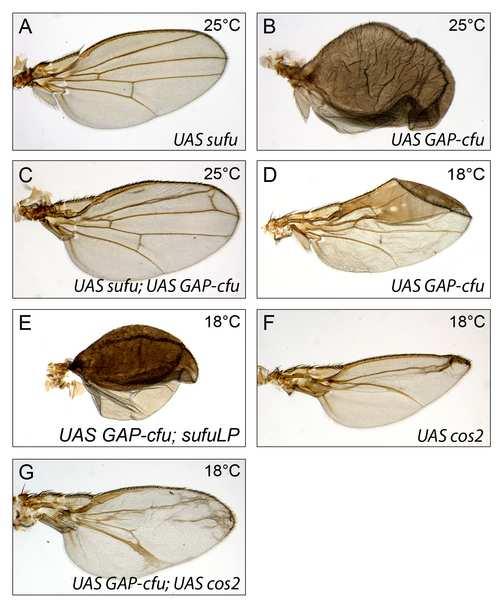
5 development (Figure 2B). By contrast, GAP-CFP-FU gave a very strong wing phenotype characteristic of strong ectopic HH pathway activation (Figure 2C, D and data not shown). In the wing disc, HH emanating from the posterior cells stabilizes full-length CI (CI-FL) [16] and induces the differential transcription of target genes in anterior cells near the anteroposterior boundary [10, 12, 17]. Consistent with its phenotypic effects, GAP-CFP-FU (but not GFP-FU) overproduction in the wing disc led to ectopic anterior (i) accumulation of CI-FL (Figure 2E and H) and (ii) expression of all the HH-responsive genes tested, including those known to respond to the highest levels of HH, including ptc itself, collier (col) and engrailed (en) (Figure 2F-L ). In fu- mutants, the production of GAP-CFP-FU rescued the (fu - ) phenotype and even led to overactivation of the pathway at the antero-posterior border (Figure 2M-N and Supplemental Data Figure S2), thus excluding the possibility that GAP-CFP-FU acted by sequestering a negative regulator of the endogenous FU protein. In contrast, the overproduction of COS2 or SU(FU), two known antagonists that interact physically with FU [18, 19], strongly decreased the activating effects of GAP-CFP-FU (Supplemental Data Figure S3). Accordingly, the loss of SU(FU) function led to an aggravation of the phenotypes induced by GAP-CFP-FU. The loss of one copy of cos2 had no effect (data not shown). In conclusion, the overproduction of a form of FU anchored to the membrane is sufficient to induce medium- to high-level activation of the HH pathway in the wing disc, raising the possibility that the subcellular distribution of FU plays a key role in controlling its activity. The wild-type and membrane-tethered forms of FU control the subcellular distribution, stability and phosphorylation of SMO 4
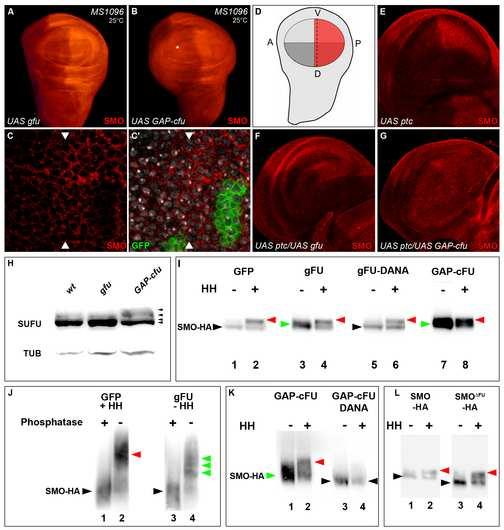
6 Surprisingly, the loss of one dose of the smo gene (flies heterozygous for the amorphic smo D16 allele), [4, 20] had an epistatic effect on the activating effects of GAP-CFP-FU, restoring an almost wild-type wing phenotype and normal levels of PTC accumulation (Figure 2O-O and P-P ). This is the first report of a dominant effect of a loss-of-function allele of smo and argues for of a dose-dependent effect of smo. SMO is stabilized in response to HH in the posterior compartment and in the anterior cells close to the antero-posterior boundary [21]. In GAP-CFP-FU-producing discs (but not in GFP-FU-producing discs), SMO accumulated throughout the disc, even in the anterior region, which contains no HH (Figure 3A, B and D). We also induced clones of cells homozygous for fu Z4, a deficiency including the fu gene [22]. In the posterior compartment, where high levels of HH normally stabilize SMO, fu Z4 /fu Z4 clones displayed low levels of endogenous SMO protein accumulation (Figure 3C and C ). These results suggest that FU is required in the wing imaginal disc for full SMO stabilization in cells receiving the HH signal and that, in the absence of HH, the anchorage of FU to the membrane is sufficient to increase SMO accumulation. In Cl-8 cells without HH, GAP-CFP-FU (alone or with SMO-RFP) was present at the plasma membrane and in large vesicular structures beneath the plasma membrane (Figure 1K- K and data not shown). In cotransfected cells, SMO-RFP, which is normally vesicular in the absence of HH, was present at the plasma membrane where it almost completely colocalized with GAP-CFP-FU (Figure1K-K ). In contrast, the distribution of SMO FU -RFP was not affected by GAP-CFP-FU (Figure 1L-L ), indicating that GAP-CFP-FU recruited SMO-RFP by interacting with it. In similar conditions (without HH), RFP-COS did not colocalize with SMO-GFP (Supplementary Data Figure S3 in [9]), and GAP-CFP-COS2 neither interacted with SMO-RFP nor relocalized it, despite being present at the plasma membrane 5
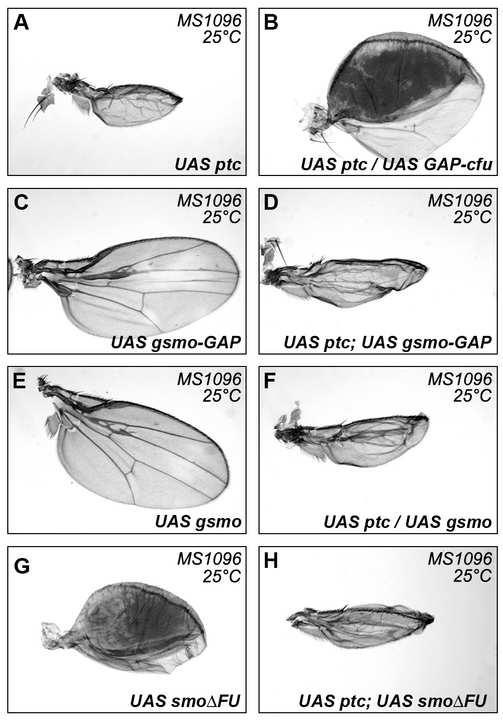
7 (Supplemental Data Figure S4). Both these data argue against a potential bridging effect of endogenous COS2. In conclusion, (i) the constitutive activation of the HH pathway induced by GAP-CFP-FU expression requires endogenous SMO, (ii) GAP-CFP-FU can recruit SMO (but not SMO FU - RFP) to the membrane of Cl-8 cells (iii) GAP-CFP-FU and FU stabilize SMO in vivo. Thus, FU acts, directly or indirectly, on SMO. The activating effects of FU anchored to the membrane cannot be inhibited by ptc overexpression PTC has a negative effect on SMO and its overproduction inhibits the HH pathway [21, 23] (Figure 2Q). However, PTC overproduction was unable to attenuate the effects of GAP-CFP- FU (Figure 2 R). In contrast, PTC overproduction was able to counter the activating effects of GFP-SMO-GAP or SMO FU, which activate the HH pathway at low and high levels, respectively (Supplemental Data Figure S5). When ptc was expressed alone with the MS1096 driver, which gives strong expression in the dorsal compartment of the wing pouch [24] (Supplemental Data Figure S6), much lower levels of posterior SMO accumulation were observed in the dorsal compartment (Figure 3E). Coexpression of the GAP-CFP-FU construct, but not of GFP-FU, (Figure 3F and G) abolished this effect of PTC, as it resulted in uniform SMO accumulation. This result is consistent with a previous report [25] showing that the increase in SMO accumulation in response to HH observed when PTC levels are reduced (by RNA interference) is abolished by a decrease in FU levels. Thus, our data demonstrate that the activation and stabilization of SMO by GAP-CFP-FU cannot be downregulated by PTC. 6
8 Wild-type FU and FU anchored to the membrane modulate the phosphorylation of SU(FU) and SMO FU is required for the phosphorylation of SU(FU) [25]. We investigated the effects of GAP-CFP-FU on SU(FU) phosphorylation, by western blot analysis of imaginal wing disc extracts (Figure 3H). In wild-type discs and in discs expressing GFP-FU, a small fraction of SU(FU) was phosphorylated, probably due to the presence of endogenous HH. The phosphorylated SU(FU) fraction was more abundant in discs producing GAP-CFP-FU (driven with MS1096). Thus, anchoring FU to the membrane can promote the phosphorylation of SU(FU). SMO is hyperphosphorylated in response to HH. This phosphorylation involves the campdependent protein kinase (PKA) and casein kinase I (CKI) and is necessary for the activation of SMO [26-28]. We monitored the phosphorylation of SMO tagged with HA (SMO-HA) in extracts of Drosophila S2 cells (Figure 3I). In the absence of HH, GFP-FU led to the appearance of slowly migrating forms of SMO-HA (green arrowhead), at intermediate positions between the unphosphorylated form of SMO-HA (black arrowhead) and the hyperphosphorylated forms induced by HH (red arrowhead). These intermediate forms corresponded to phosphorylated isoforms of SMO-HA, as they disappeared after incubation of the extracts with alkaline phosphatase (Figure 3J). GAP-CFP-FU had a similar effect on SMO-HA phosphorylation (Figure 3I). GAP-CFP-FU also reproducibly stabilized SMO-HA (see Figure 3 and for supplemental controls the Supplemental Data Figure S7), resulting in larger amounts of phosphorylated SMO-HA than obtained with GFP-FU. Thus, both GFP-FU and GAP-CFP-FU can induce the phosphorylation of SMO, and our data suggest that FU may activate the pathway by increasing the amount of phosphorylated SMO accumulated. 7
9 The kinase activity of FU is involved both in the phosphorylation of SMO and the activating effects of GAP-CFP-FU We investigated whether the phosphorylation of SMO-HA induced by FU was dependent on the kinase activity of FU, by testing two mutants of FU (Figure 2A): GFP-FU-DANA, mutated for two amino acids crucial for the phospho-transfer reaction, and GFP-FU-AS, in which a conserved residue (Thr 158) potentially involved in activating the autophosphorylation of FU was modified. Both mutants were shown to have lost their ability to enhance the transcriptional response of an HH-sensitive reporter gene in S2 cells [29]. We found that GFP-FU-DANA induced no phosphorylation of SMO-HA (Figure 3I), whereas FU-AS induced only very low levels of phosphorylation (data not shown). Similar results were obtained with GAP-CFP-FU-DANA (Figure 3K) and GAP-CFP-FU-AS (data not shown). However, these two proteins were present at the plasma membrane, where they recruited SMO-RFP (Supplemental Data Figure S8). Thus, neither its kinase activity nor its Thr 158 are required for the interaction of GAP-CFP-FU with SMO or the recruitment of SMO at the plasma membrane but they are required to promote SMO phosphorylation. FU-GFP-DANA expression unexpectedly induced a [fu ]-like phenotype and FU-GFP-AS induced mild downregulation of the HH pathway (Supplemental Data Figure S8). GAP-CFP- FU-AS and GAP-CFP-FU-DANA gave similar phenotypes, indicating that the negative effects of FU-DANA/AS could not be overcome by tethering these molecules to the membrane. As these mutants of CFP-FU, (with or without their GAP anchor) recruit FU-GFP to the plasma membrane (Supplemental Data Figure S9), they may exert their dominant negative effects by dimerizing with endogenous FU. We detected no effect of GFP-FU or GAP-CFP-FU on the phosphorylation of SMO-HA induced by HH. Nevertheless, GAP-CFP-FU-DANA (and to a lesser extent, GFP-FU-DANA) 8
10 significantly decreased the fraction of SMO-HA phosphorylated in response to HH (Figure 3K and data not shown). Moreover, SMO FU -GFP, which cannot interact with FU, displayed a lower fraction of phosphorylated isoforms in response to HH than SMO (Figure 3L and Supplemental Data Figure S10). Thus, both the kinase activity of FU and the interaction between FU and SMO are required for full SMO phosphorylation in response to HH. DISCUSSION This work provides new information about (i) the mechanisms by which the activation of SMO is transduced to its cytoplasmic effector FU, (ii) the mechanisms of FU activation, and (iii) a novel positive feedback loop between FU and SMO. Here and in a previous study [9], we provide evidence that SMO controls the subcellular distribution of two of its physical partners, FU and COS2, recruiting them to the plasma membrane in response to HH. Our data also suggest that FU may link COS2 to SMO in a vesicle-associated complex in the absence of HH, whereas FU and COS2 may independently bind SMO at the plasma membrane in the presence of HH. Thus, HH may not only promote, via SMO, the recruitment of FU and COS2 to the plasma membrane, it may also modulate the nature of interactions between these three proteins. Several non exclusive mechanisms seem to be involved in controlling FU activity. First, the forced localisation of FU at the membrane induces strong SMO-dependent activation of the pathway in the wing. This study is the first to report a dominant active form of this type of kinase. Second, we show here that the presence of a conserved Thr in the activating loop is important for the promotion of full SMO phosphorylation and for the activating effects of GAP-FU. Thus, as FU is known to be phosphorylated in response to HH [30], the phosphorylation of this loop by auto-phosphorylation or by other kinases may be a key 9
11 element in FU regulation. Third, HH may regulate FU by controlling its dimerization or its interaction with potential regulatory proteins. Possible FU dimerization is consistent with (i) the reported interaction between the regulatory domain of FU and its kinase domain [31], (ii) the recruitment to the plasma membrane of wild-type FU by the wild-type and mutant forms of GAP-FU (Supplemental Data, Figure S8), and (iii) the dominant negative effects of FU mutants with modified kinase domains (see Supplemental Data Figure S2 and 8). We present here evidence for of a novel, positive feedback loop in which SMO and FU enhance each other's activities. Indeed, SMO promotes the relocalization of FU to the plasma membrane and is required for the activating effects of GAP-FU, whereas both GAP- FU and FU control SMO stability and phosphorylation. FU kinase activity is required for SMO phosphorylation and for the activating effects of GAP-FU, but not for its association with SMO. FU may phosphorylate SMO directly or may act on other substrates, indirectly facilitating SMO phosphorylation, inhibiting phosphatases or stabilizing phosphorylated SMO. Both FU activity and its interaction with SMO seem to be required for full hyperphosphorylation of SMO in response to HH. In the wing disc, FU is required principally for responses to the highest levels of HH present at the antero-posterior border [10-12], where SMO is active despite the strong upregulation of ptc. The effects of GAP-FU and FU on SMO reported here provide the first clues to a putative mechanism (FU-dependent phosphorylation and stabilization of SMO), potentially accounting for the resistance of SMO to the high level of PTC induced by HH in responding cells. We propose the following model: (i) The HH-induced relocalization of SMO to the plasma membrane leads to the recruitment of FU and COS2 at this membrane. (ii) FU, in turn, acts on SMO, probably by further enhancing its phosphorylation, to stabilize it further and prevent its inhibition by PTC. It is not yet possible to determine whether FU regulates SMO directly or 10
12 indirectly. The kinesin COS2 may be also part of this regulatory loop. (iii) The stabilized, activated SMO/FU/COS2 complex at the plasma membrane then promotes the accumulation and activation of CI-FL, leading to the activation of HH target genes, including ptc. We previously reported that SMO FU, which does not bind FU, is constitutively active, suggesting that FU may also act as a negative regulator of SMO in the absence of the HH signal [9]. Thus, FU may act as a switch, sensing the level of HH, inhibiting SMO in the absence of HH or activating the pathway in response to high levels of HH. Interestingly, the existence of such regulatory loops may account for the bistability properties of signaling pathways and explain how graded levels of signal may act as morphogens, leading to differential cell responses (for review see [32]). In conclusion, we found that FU was recruited by SMO at the plasma membrane in response to HH and that this recruitment was directly dependent on the physical interaction of FU with SMO. We also found that the expression of a membrane-anchored form of FU (GAP- FU) constitutively activated the HH pathway, indicating that FU activity may be regulated by its subcellular location. Surprisingly, the activating effects of GAP-FU require a wild-type dose of endogenous SMO. We also report evidence that (i) FU and GAP-FU induce the phosphorylation of SMO, (ii) GAP-FU recruits SMO to the plasma membrane, (iii) GAP-FU renders SMO resistant to the destabilizing effects of PTC and (iv) FU controls the level of accumulation of SMO in the wing imaginal disc. Thus, our data demonstrate that FU, which is generally considered to be an effector of SMO, can also act on SMO. EXPERIMENTAL PROCEDURES Plasmids 11
13 All expression vectors were constructed by the Gateway recombination method (Invitrogen). The constructs used here are presented in the supplemental data. Cell culture and transfection Cl-8 cells were cultured as previously described [33]. Cells were plated on concanavalin A-coated coverslips and incubated for 24 h. Transient transfections were then carried out using FlyFectin (OZ bioscience). S2 cells were cultured in Schneider medium and transfected using Effectene reagent (Qiagen). Before observation, cells were fixed by incubation for 30 minutes with 4% paraformaldehyde. Imaginal disc immunostaining and imaging (see also supplemental data). The primary antibodies used were: mouse anti-ptc, 1/100 [24], mouse anti-en, 1/1000 (4D9, from DSHB [34]), mouse anti-smo, 1/1000 (20C6, from DSHB, [25]), rat anti-ci, 1/5 (2A1, [35]), rabbit anti-col, 1/250 [11] and rabbit polyclonal anti- -galactosidase, 1/1000 (from ICN/Cappel). Secondary antibodies were obtained from Jackson ImmunoResearch Laboratory and were all used at a dilution of 1/200. All the images of wing imaginal discs presented were acquired with a Leica-SP2-AOBS microscope except when indicated otherwise. Western blotting Proteins were extracted from wing discs by the NaOH/TCA method [36] and from transfected S2 cells, using RIPA buffer. Protein extracts corresponding to 20 wing discs or to 30 g of S2 cells extracts were fractionated by SDS-PAGE and transferred to a nitrocellulose membrane (Schleicher & Schuell). Blots were probed with a polyclonal rabbit antibody specific for SU(FU) [19], a mouse monoclonal antibody specific for TUB (Sigma) or a rat monoclonal antibody against HA (Roche). Bound antibody was detected with secondary antibody conjugated with horseradish peroxidase (Biovalley Vector Lab.). The 12
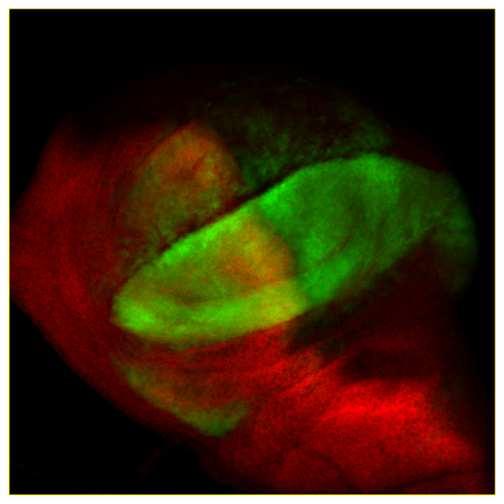
14 immunolabeled bands were detected with the enhanced chemiluminescence detection system (Supersignal West Pico Chemiluminescent Substrate, Pierce). ACKNOWLEDGMENTS We thank S. Cohen, I. Guerrero, J, Jiang, L. Ruel, F. Schweisguth, P. Thérond, C. Tong, M. van den Heuvel, A-M Voie and A. Zhu for providing plasmids, antibodies, fly strains, cell lines and technical advice, S. Malpel for his participation in the construction of the GAP-CFP- FU vector and of the transgenic UAS-GAP-CFP-fu flies, R. Benkiran for her participation in some experiments, A. Brigui, C. Chamot, M. Coppey, R. Karess, C. Lamour-Isnard, T. Piolot, F. Schweisguth, M. Tramier and H. Tricoire for their help and support. We also thank the Cell Press editors and anonymous reviewers for their suggestions. The monoclonal antibodies obtained from the Developmental Studies Hybridoma bank were developed under the auspices of the NICHD and maintained by the University of Iowa. Drosophila embryo injections were carried out by BestGene Inc Service. English usage was corrected by Alex Edelman & Associates. This work was supported by the CNRS, University Paris 7 and ARC (grant no. 4797), by an ACI Biologie du développement et physiologie integrative (grant no ). The imaging facilities were partially funded by the ARC, the region Ile de France (SESAME) and the University Paris VII. S. C. was ATER at the University Paris 7, and was also supported by Fondation des Treilles. LEGENDS Figure 1: Subcellular distribution of SMO and FU with and without HH. Confocal images of transfected Cl-8 cells expressing fluorescent tagged versions of FU and/or SMO without HH (A, C, E, G-G, I-I, K-K and L-L ) or with HH (B, D, F, H-H and J-J ). 13
15 (A-B) Transfected cells producing GFP-FU (noted FU, in red), (C-D) SMO-GFP (SMO, in green) or (E-F) SMO FU -GFP (SMO FU, in green). (G-J) Cotransfected cells producing RFP-FU (FU, in red) with SMO-GFP (green) (-HH in G-G and +HH in H-H ), or SMO FU -GFP (SMO FU, in green) (-HH in G-G and +HH in J- J ); (K-L) Cotransfected cells producing GAP-CFP-FU (GAP-FU, in green) with SMO-RFP (SMO, in red) (K-K ) or SMO FU -RFP (red) (L-L ) in the absence of HH. Merged pictures are shown in G -L. Arrow in G : vesicular structures. Arrowhead in H : plasma membrane. Figure 2: Activation of the HH pathway by a form of FU anchored at the plasma membrane. (A) Schematic representation of the FU constructs used in this study. FU kinase domain (Kin, aa 1-255, in pale gray), FU regulatory domain (Reg, aa , in dark gray). FU- DANA has two point mutations affecting the phosphor-transfer reaction. In FU-AS, Thr158 is replaced by Ala (green triangle). GAP43 domain (yellow box), fluorescent protein : GFP in green or CFP in blue. (B-D) Wing phenotypes of flies (or imago in C) overexpressing (B) MS1096; UAS-GFP-fu (indicated as gfu); (C) MS1096; UAS-GAP-CFP-fu (indicated as GAP-cfu); (D) UAS-GAP- CFP -fu; 71B. 71B is a weaker driver than MS1096, both are expressed in the wing pouch, to similar levels, in the anterior and posterior compartments (See also in the Supplemental data the experimental procedures section and the Figure S6). A more detailed legend to this figure is also given in the Supplemental data. (E-L) Effects of GFP-FU alone (E-G) or fused to a GAP- anchor (GAP-CFP-FU in H, I, J- L) overproduction in wing discs under control of MS1096 (E-J) or in clones (K, L). (E, H) CI- 14
16 FL immunolabeling, (F, I) dpp expression, as reported by a dpp-lacz construct (nuclear - galactosidase immunostaining, in blue, with EN immunostaining to visualize the posterior compartment, in red); (G, J) PTC immunodetection. (See also supplemental legend); (K-L) anterior clones (labeled with GFP, in green) of GAP-CFP-FU-expressing cells with en (red immunostaining in K) and col (red immunostaining in L) expression. In (K), the normal anterior expression of en along the A/P border is indicated by (*). The effects of GAP-CFP- FU could be seen in all anterior clones, even away from the boundary, indicating that they occurred independently of HH. (M-N) fu M1 /Y; 71B (M) and fu M1 /Y; UAS-GAP-CFP-fu; 71B (N) wings. The fusion of the longitudinal veins (LV) 3 and 4 and narrowing of the LV3-4 intervein region are characteristic of fu mutants. The effect on GAP-cFU varies with the fu allele used (see Supplemental Data, Figure S2). (O-P) Wing phenotype (O and P) and immunodetection of PTC (O and P ) in smo + /smo + (O and O ) or smo D16 /smo + (P and P ) flies expressing UAS-GAP-CFP-fu under control of the MS1096 driver. (Q-R) Wings of flies overexpressing UAS-ptc (Q); UAS-ptc and UAS-GAP- CFP-fu (R) under MS1096 driver; Wings in B were obtained from dissected imagos as the adults were unable to emerge. All wing discs in this study are oriented with the anterior to the left and the dorsal part to the bottom and the wings are shown with the anterior to the top and the proximal end to the left. 1 to 4: LV1 to LV4. All pictures are confocal sections, except for (E, G, H and J), which are epifluorescence images. 15
17 Figure 3: GAP-CFP-FU and FU modulate the levels of SMO accumulation and phosphorylation (A-D) Immunostaining of endogenous SMO (red).wing imaginal discs expressing the GFP-FU (A) or GAP-CFP-FU (B) construct. The asterisk in B indicates the ectopic accumulation of SMO in the anterior region. Wing disc with a posterior fu Z4 /fu Z4 clone (labeled with GFP in C, green) (C). In the anterior region, the GFP-cells are fu Z4 /fu + and the already low level of SMO would make it difficult to see any further reduction. Nuclei are shown in gray (C ). Note that only a few (8 to 10) rows of anterior cells are shown here and that in this region, HH emanating from the P cells is still present, leading to a gradual decrease in SMO accumulation. The difference between A and P cells is therefore more visible when comparing the P cells (outside the clone) with the most anterior cells. (D) Schematic representation of a wing imaginal disc showing both the stronger accumulation of SMO (red) in the P compartment and in A near the A/P boundary (dotted line) and the pattern of MS1096-driven expression, which is stronger in the dorsal compartment (dark gray stripes) than in the ventral compartment (light gray stripes) [24]. A/P: antero-posterior axis; D/V: dorso-ventral axis. See also the pattern of GFP MS1096-driven expression in Supplemental Data, Figure S6. (E-G) Immunostaining of endogenous SMO in wing imaginal discs expressing UAS-ptc (E); UAS-ptc and UAS-GFP-fu (F) or UAS-ptc and UAS-GAP- CFP-fu (G). In this figure, the expression of all transgenes was driven by MS1096 at 25 C. (H) Immunodetection of endogenous SU(FU) in extracts of MS1096 (wt), MS1096; UAS- GFP-fu (gfu) or MS1096; UAS-GAP-CFP-fu (GAP-cfu) wing imaginal discs. Tubulin (TUB). The black arrowheads indicate the different SU(FU) isoforms. (I) Immunodetection of SMO-HA in extracts of S2 cells transiently expressing SMO-HA with GFP as a control (1-2) or along with GFP-FU (3-4) GFP-FU-DANA (5-6) or GAP-CFP- 16
18 FU (7-8) without (-) (odd-numbered lanes) or with (+) (even-numbered lanes) HH. See also Figure S7 for the transfection and loading controls. (J) Immunodetection of SMO-HA in the extracts analyzed in lanes 2 and 3 of panel B. Samples in lanes 1 and 3 of this panel were treated with alkaline phosphatase, but not samples in lanes 2 and 4. Here, the gel was run for a longer period than in panel (B). (K) Immunodetection of SMO-HA in extracts of S2 cells transiently expressing SMO-HA with GAP-CFP-FU (1-2) or with GAP-CFP-FU-DANA (3-4). (L) Immunodetection of SMO-HA in extracts of S2 cells transiently expressing SMO (1-2) or SMO FU -HA (3-4) without (1, 3) or with (2, 4) HH. For quantification of the phosphorylation levels see Supplemental Figure S10. Note the increased levels of SMO FU - HA accumulation which can be correlated with its reported increased activity [9]. REFERENCES 1. Beachy, P.A., Karhadkar, S.S., and Berman, D.M. (2004). Tissue repair and stem cell renewal in carcinogenesis. Nature 432, Lau, J., Kawahira, H., and Hebrok, M. (2006). Hedgehog signaling in pancreas development and disease. Cell Mol Life Sci 63, Nakano, Y., Guerrero, I., Hidalgo, A., Taylor, A., Whittle, J.R.S., and Ingham, P.W. (1989). A protein with several possible membrane-spanning domains encoded by the Drosophila segment polarity gene patched. Nature 341, van den Heuvel, M., and Ingham, P.W. (1996). smoothened encodes a receptor-like serpentin protein required for hedgehog signalling. Nature 382, Alcedo, J., and Noll, M. (1997). Hedgehog and its patched-smoothened receptor complex: a novel signalling mechanism at the cell surface. Biol Chem 378, Hooper, J.E., and Scott, M.P. (2005). Communicating with Hedgehogs. Nat Rev Mol Cell Biol 6, Robbins, D.J., Nybakken, K.E., Kobayashi, R., Sisson, J.C., Bishop, J.M., and Therond, P.P. (1997). Hedgehog elicits signal transduction by means of a large complex containing the kinesin-related protein costal2. Cell 90, Sisson, J.C., Ho, K.S., Suyama, K., and Scott, M.P. (1997). Costal2, a novel kinesinrelated protein in the Hedgehog signaling pathway. Cell 90, Malpel, S., Claret, S., Sanial, M., Brigui, A., Piolot, T., Daviet, L., Martin-Lanneree, S., and Plessis, A. (2007). The last 59 amino acids of Smoothened cytoplasmic tail directly bind the protein kinase Fused and negatively regulate the Hedgehog pathway. Dev Biol 303,
19 10. Sanchez-Herrero, E., Couso, J.P., Capdevila, J., and Guerrero, I. (1996). The fu gene discriminates between pathways to control dpp expression in Drosophila imaginal discs. Mech Dev 55, Crozatier, M., Glise, B., and Vincent, A. (2002). Connecting Hh, Dpp and EGF signalling in patterning of the Drosophila wing; the pivotal role of collier/knot in the AP organiser. Development 129, Alves, G., Limbourg, B.B., Tricoire, H., Brissard, Z.J., Lamour, I.C., and Busson, D. (1998). Modulation of Hedgehog target gene expression by the Fused serinethreonine kinase in wing imaginal discs. Mech Dev 78, Strittmatter, S.M., Igarashi, M., and Fishman, M.C. (1994). GAP-43 amino terminal peptides modulate growth cone morphology and neurite outgrowth. J Neurosci 14, Zhu, A.J., Zheng, L., Suyama, K., and Scott, M.P. (2003). Altered localization of Drosophila Smoothened protein activates Hedgehog signal transduction. Genes Dev 17, Ascano, M., Jr., Nybakken, K.E., Sosinski, J., Stegman, M.A., and Robbins, D.J. (2002). The carboxyl-terminal domain of the protein kinase fused can function as a dominant inhibitor of hedgehog signaling. Mol Cell Biol 22, Aza, B.P., Ramirez, W.F., Laget, M.P., Schwartz, C., and Kornberg, T.B. (1997). Proteolysis that is inhibited by hedgehog targets Cubitus interruptus protein to the nucleus and converts it to a repressor. Cell 89, Ohlmeyer, J.T., and Kalderon, D. (1998). Hedgehog stimulates maturation of Cubitus interruptus into a labile transcriptional activator. Nature 396, Monnier, V., Ho, K.S., Sanial, M., Scott, M.P., and Plessis, A. (2002). Hedgehog signal transduction proteins: contacts of the Fused kinase and Ci transcription factor with the kinesin-related protein Costal2. BMC Dev Biol 2, Monnier, V., Dussillol, F., Alves, G., Lamour, I.C., and Plessis, A. (1998). Suppressor of fused links fused and Cubitus interruptus on the hedgehog signalling pathway. Curr Biol 8, Chen, Y., and Struhl, G. (1996). Dual roles for patched in sequestering and transducinghedgehog. Cell 87, Denef, N., Neubuser, D., Perez, L., and Cohen, S.M. (2000). Hedgehog induces opposite changes in turnover and subcellular localization of patched and smoothened. Cell 102, Preat, T., Therond, P., Lamour, I.C., Limbourg, B.B., Tricoire, H., Erk, I., Mariol, M.C., and Busson, D. (1990). A putative serine/threonine protein kinase encoded by the segment- polarity fused gene of Drosophila. Nature 347, Johnson, L.N., Noble, M.E., and Owen, D.J. (1996). Active and inactive protein kinases: structural basis for regulation. Cell 85, Capdevila, J., Pariente, F., Sampedro, J., Alonso, J.L., and Guerrero, I. (1994). Subcellular localization of the segment polarity protein patched suggests an interaction with the wingless reception complex in Drosophila embryos. Development 120, Lum, L., Zhang, C., Oh, S., Mann, R.K., von Kessler, D.P., Taipale, J., Weis-Garcia, F., Gong, R., Wang, B., and Beachy, P.A. (2003). Hedgehog signal transduction via Smoothened association with a cytoplasmic complex scaffolded by the atypical kinesin, Costal-2. Mol Cell 12, Apionishev, S., Katanayeva, N.M., Marks, S.A., Kalderon, D., and Tomlinson, A. (2005). Drosophila Smoothened phosphorylation sites essential for Hedgehog signal transduction. Nat Cell Biol 7,
20 27. Jia, J., Tong, C., Wang, B., Luo, L., and Jiang, J. (2004). Hedgehog signalling activity of Smoothened requires phosphorylation by protein kinase A and casein kinase I. Nature 432, Zhang, C., Williams, E.H., Guo, Y., Lum, L., and Beachy, P.A. (2004). Extensive phosphorylation of Smoothened in Hedgehog pathway activation. Proc Natl Acad Sci U S A 101, Fukumoto, T., Watanabe-Fukunaga, R., Fujisawa, K., Nagata, S., and Fukunaga, R. (2001). The fused protein kinase regulates Hedgehog-stimulated transcriptional activation in Drosophila Schneider 2 cells. J Biol Chem 276, Therond, P.P., Knight, J.D., Kornberg, T.B., and Bishop, J.M. (1996). Phosphorylation of the fused protein kinase in response to signaling from hedgehog. Proc Natl Acad Sci U S A 93, Ascano, M., Jr., and Robbins, D.J. (2004). An intramolecular association between two domains of the protein kinase Fused is necessary for Hedgehog signaling. Mol Cell Biol 24, Lander, A.D. (2007). Morpheus unbound: reimagining the morphogen gradient. Cell 128, van Leeuwen, F., Samos, C.H., and Nusse, R. (1994). Biological activity of soluble wingless protein in cultured Drosophila imaginal disc cells. Nature 368, Patel, N.H., Martin-Blanco, E., Coleman, K.G., Poole, S.J., Ellis, M.C., Kornberg, T.B., and Goodman, C.S. (1989). Expression of engrailed proteins in arthropods, annelids, and chordates. Cell 58, Motzny, C.K., and Holmgren, R. (1995). The Drosophila cubitus interruptus protein and its role in the wingless and hedgehog signal transduction pathways. Mech. Dev. 52, Riezman, H., Hase, T., van Loon, A.P., Grivell, L.A., Suda, K., and Schatz, G. (1983). Import of proteins into mitochondria: a 70 kilodalton outer membrane protein with a large carboxy-terminal deletion is still transported to the outer membrane. Embo J 2,
21 Figure 1
22 Figure 2
23 Figure 3
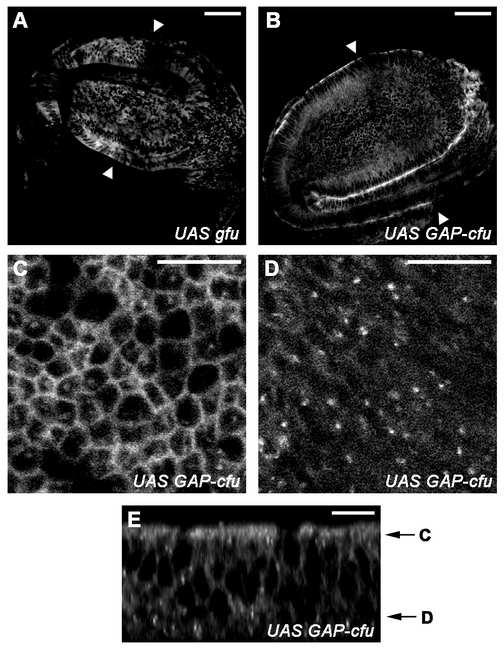
24 Supplemental Data text SUPPLEMENTAL DATA Supplemental legends to Figure 2: (B-D): The MS1096; UAS-GAP-cfu flies were unable to emerge from pupae, probably due to their enlarged and inflated wings. Imago from MS1096; UAS-GAP-cfu dissected pupae (C) present wings with the A compartment invaded by ectopic vein tissue associated with supernumerary campaniform sensillae, which are specific to regions controlled by high levels of HH (Figure 2C and data not shown). d1 and d2 are the distances measured with ImageJ software. The average ratio d2/d1 of flies overexpressing or not over-expressing the gfu and GAP-cFU constructs under control of the 71B driver shows an increase in LV3/LV4 intervein tissue: d2/d1 = 0.22 ± 0.01 for WT wings, 0.22 ±0.01 for UAS-gfu wings and 0.25 ± 0.04 for UAS-GAP-cfu wings. Note that GAP-cFU had no effect on the innervated double-row bristles. (K-L): These effects were observed in all anterior clones in the wing pouch, even away from the boundary, indicating that they occurred independently of HH. These effects are only observed within the clone limits. Posterior clones have no effect (data not shown). Supplemental Figures: Figure S1: Accumulation and subcellular localization of GAP-CFP-FU in the wing imaginal disc As reported for endogenous FU, overexpressed GFP-FU (A) and GAP-CFP-FU (B) are more abundant in the A compartment (where there is little or no HH) than in the P compartment. Although we cannot conclude that GAP-cFU is also destabilized by the HH signal, it does appear to be sensitive. (C and D) are magnified views of apical and basolateral confocal sections, respectively, of a wing imaginal disc expressing GAP-cFU. (E) A Xz view reconstruction of the wing pouch across the A/P axis. GAP-FU is localized principally at the plasma membrane on the apical side of the disc, and is also present in the vesicular structure below. The expression of GFP-FU and GAP- CFP-FU was driven by MS1096 at 25 C, a driver equally expressed in the anterior and posterior compartments (See Figure S6). The A/P axis is indicated by two arrowheads, in A to D, with A on the left. In E, the apical side is up. In (A) and (B), scale bars correspond to 50 m, and in (C-E), scale bars correspond to 10 m. Xz view reconstructions were obtained using ImageJ
25 software (Rasband, W.S., ImageJ, U. S. National Institutes of Health, Bethesda, Maryland, USA, ). Figure S2: Effects of fu - mutations on the activating effects of GAP-CFP-FU (A) Wild-type wing. (B) Wing overexpressing the GAP-CFP-FU construct, showing widening of the LV3/LV4 intervein and a thickening of LV3. fu 1 /Y (C), fu RX2 /Y (E) and fu M1 /Y (G) wings. Overexpression of the GAP-CFP-fu construct in fu 1 /Y (D), fu RX2 /Y (F) and fu M1 /Y (H). Overexpression was driven by 71B and wings from male insects were observed. In all cases tested, expression of the GAP-CFP-FU construct (driven by 71B, [1]) in the wing pouch, rescued the fu - phenotype. GAP-CFP-FU even overactivated the pathway at the antero-posterior border, as shown by the widening of the LV3/LV4 intervein region and the expansion of LV3 tissue. The effects of GAP- CFP-FU were not affected in the fu RX2 mutant, but they were attenuated, although not abolished in fu 1 and fu M1 mutants. Thus, in the presence of high levels of HH (near the antero-posterior border) the activating effects of GAP-cFU did not require the kinase activity of endogenous FU. This indicates that the activating effects of GAP-FU do not simply result from the activation of endogenous FU. However, in the presence of only small amounts of HH and in the absence of HH, the activation by GAP-CFP-FU was reduced by the presence of an endogenous FU protein with an altered kinase domain. There are several possible explanations for this, including a dominant negative effect of endogenous FU with a mutant kinase domain. When modeling the structure of the kinase domain of FU (data not shown), the first 80 aa of FU, which are present in fu MI ) correspond to most of its N-terminal domain, called the small lobe. The equivalent domain in other kinases has been shown to bind different partners. A striking example is provided by SNF1 kinase, a protein kinase closely related to FU: this kinase can form homodimers via interactions involving its small lobe [2]. The kinase domain of FU has been reported to bind to its regulatory domain [3, 4] and to its putative target SU(FU) [5]. Thus, the first 80 aa of FU M1 may be sufficient to induce a dominant-negative effect.
26 Figure S3: GAP-cFU-dependent activation of the HH pathway is regulated by the inhibitory regulators SU(FU) and COS2 The overexpression of various UAS transgenes was driven by MS1096 at 25 C (A-C) or at 18 C (D-G) (as indicated). The wing overexpressing SU(FU) (A) had only a very weak phenotype, if at all (e.g. distal extra vein near LV3). The overexpression of SU(FU) almost abolished the phenotype induced by GAP-cFU overexpression (compare C with B). This decrease is not due to the dosage of GAL4 protein controlled by a second UAS (data not shown). The loss of Su(fu) function (in flies homozygous for the amorphic mutation, Su(fu) LP ) was associated with no visible phenotype itself (data not shown), but it exacerbated the phenotype of GAP-CFP-FU-overexpressing wings (visible at 18 C, compare E with D). The overexpression of COS2 (F) strongly inhibited the HH pathway (F) and strongly decreased the GAP-CFP-FU-dependent phenotype (compare G with D). Su(fu) LP was described in[6], UAS-cos2 was kindly provided by M. Scott [7], and UAS-Su(fu) was described in [8]. Figure S4: GAP-CFP-COS2 (in green) does not colocalize with SMO-RFP (in red) Transfected S2 cells without HH. Figure S5: The effects of PTC overproduction are not suppressed by SMO or SMO-GAP (A-H) Wings of flies overexpressing UAS-ptc (A); UAS-ptc and UAS-GAP-cfu (B); UAS-GFP-smo-GAP (C); UAS-ptc and UAS-GFP-smo-GAP (D); UAS-GFP-smo (E); UAS-ptc and UAS- GFP-smo (F); UAS-GFPsmo FU (G); UAS-ptc and UAS-GFP-smo FU (H). The overproduction of GFP-SMO and GFP-SMO-GAP induced low and medium levels of HH pathway activation, respectively [9, 10]. GFP-SMO-GAP had a stronger effect than GFP-SMO, but a much weaker effect than GAP-CFP-FU. Coexpression of the PTC construct with those encoding GFP-SMO-GAP or GFP-SMO led to a strong decrease in wing size and a loss of wing patterning. By contrast, coexpression of the PTC construct did not attenuate the effects of GAP-cFU. Thus, the activating effects of GAP-CFP-FU act epistatically on the inhibitory effects of PTC overproduction, whereas neither GFP-SMO-GAP nor GFP-SMO has such an effect. The lack of effect of PTC on GAP-CFP-FU-
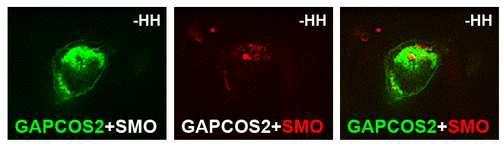
27 induced activation may be due to GAP-cFU inducing a very high level of activation. We do not favor this hypothesis because PTC has been reported to inhibit SMO substoichiometrically [11] and the overproduction of PTC led to downregulation of the effects of another very strong inducer of the HH pathway, SMO FU. Our data suggest that FU anchored to the plasma membrane does more to SMO than just recruiting it at the plasma membrane. The comparison between SMO-GAP and GAP-FU is validated by the fact that the same GAP moiety was used to anchor SMO and FU at the membrane, and therefore should act similarly on these two proteins. The GAP domain is known to be target for palmitoylation which is supposed to anchor proteins in raft domains. Noteworthy, HH has been found in lipid rafts in Drosophila, and human PTC and SMO were found in raft microdomains (Rietvel et al, J Biol Chem., 1999 and Karpen et al, J Biol Chem, 2001). Note also that several receptors of the GPCR family have been shown to be regulated by palmitoylation in response to agonists (Ross, Curr Biol, 1995, Stevens et al, J Biol Chem, 2001). However, without understanding why GFP- SMO-GAP is really activated, it is difficult to speculate. Wings in (B and G) were obtained from dissected imagos as the adults were unable to emerge. Figure S6: Pattern of MS1096 activity. MS1096, UAS-GFP imaginal disc. CI immunolabelling in red, GFP in green. Orientation is as in Figure 2. Figure S7: GFP-FU, GAP-CFP-FU and GFP-FU-DANA accumulate at comparable level. Immunodetection of FU on the blot presented in Figure 3 I. The effects of GAP-FU on the accumulation of SMO seen in the western blot presented in Figure 3I were seen in 5 independent experiments. In each experiments the same amount of proteins were loaded in all lanes. However, to ensure that the differences of effects between Gap-FU and FU or FU-DANA were not due to differences in transfection efficiency or in the stability of the different forms of FU, we checked the levels of accumulation of the various forms of FU. Figure S8: Effects of GAP-CFP-FU-DANA and GAP-CFP-FU-AS on SMO-RFP subcellular distribution and wing phenotype
28 (A-B) Cells expressing SMO-RFP (in red, A, A, B and B ) with either GAP-CFP-FU-DANA (in green, A, A ) or GAP-CFP-FU-AS (in green, B, B ). (C-F) Wings of flies overexpressing forms of FU with mutations affecting their kinase domain with or without the GAP domain (MS1096 driver at 25 C). Figure S9: Cotargeting of wild-type FU to the plasma membrane by GAP-FU Transfected Cl8 cells coexpressing different forms (WT, DANA or AS) of GAP-cFU (in green) and rfu (in red) in the absence of HH. (A-A ) Cells cotransfected with GAP-CFP-FU and RFP-FU; (B-B ) Cells cotransfected with GAP-CFP-FU- DANA and RFP-FU; (C-C ) Cells cotransfected with GAP-CFP-FU-AS and RFP-FU. In these three contexts, RFP-FU is recruited to the plasma membrane by the GAP-FU forms. These data indicate that FU can interact with itself. This cotargeting of FU forms is independent of AS and DANA mutation. Figure S10: The efficiency of SMO phosphorylation in response to HH in reduced in SMO FU. The immunoblot presented in Figure 3L was plotted using ImageJ. With SMO the HH-dependent hyperphosphorylated isoforms are always more abundant than the non phosphorylated isoform, while with SMO FU the both isoforms are equally abundant. Supplemental experimental procedures: Vectors: The full coding sequences (without the termination codon) of smo and fu cdnas were amplified by PCR, and inserted into pentr/d-topo by directional TOPO cloning. The RFP-FU, SMO-GFP and SMO FU -GFP expression vectors have been described elsewhere [9]. Rescue experiments demonstrated the functionality of the rfu and SMOg fusion proteins in Drosophila [9]. AS and DANA FU mutants were amplified by PCR from pda-flag-fu-as or pda-flag-fu-dana [12]. The DANA mutant has two alanine residues replacing Asp-125 and Asn-130, both of which are invariantly conserved among members of the protein kinase family and are involved in the phospho-transfer reaction. The AS mutant has an alanine residue
29 replacing the conserved Thr-158. The destination vectors pawr (pact5c-gw-mrfp) (where GW is the recombination cassette) were obtained from T. Murphy. The puast-nterm-m6gfp and puast-cfp-gw vectors were a gift from A. Brand. A sequence encoding the GAP peptide (MLCCMRRTKQVEKNDEDEDQKI) was inserted in frame with the CFP into the BglII site located just upstream from the CFP in puast-cfp-gw. The HH expression vector, pda-fl Hh, was a gift from D.J. Robbins. All cloning steps were performed according to the manufacturer's instructions (Invitrogen). Fly strains and genetics Flies were raised at 25 C unless otherwise specified. Mutants and transgenic strains are described in the Supplemental data. Transgenic, UAS-GFP-fu and UAS-GAP-CFP-fu strains, with or without DANA and AS mutations, were obtained by a standard P-element-mediated procedure. For clonal overexpression, hsp-flp;+/+; tub1>cd2>gal4, UAS-GFP/UAS-GFP-fu or puas-gap-cfp-fu first instar larvae were subjected to heat shock for one hour at 38 C. Mosaic analysis with a repressible cell marker (MARCM) [13]) was carried out in larvae from crosses between FRT19A, Tub-Gal80, hsp-flp; UAS-LacZ, UAS-CD8-GFP; TubGal4/TM6b females (a gift from S. Bray) and FRT19A fuz4/y males. MS1096 (chr. X, [14]), 71B (chr. III, [15]), hsp-flp; +/+; tub1>cd2>gal4, UAS-GFP /TM3 [16]. fu 1, fu M1 and fu RX2 were described by [17]. UAS-ptc/CyO and UAS-GFP-smoGAP/TM3 flies were provided by M. Scott; the smo D16 allele was provided by Van den Heuvel and is thought to be a probable null allele [18]. Analysis of wings Wings were dissected in 70% ethanol, mounted in Hoyers medium and examined for their vein and wing margin bristle phenotypes. Using ImageJ software, we measured both the width of the LV3-LV4 intervein (d1) and that of the entire wing (d2), and then calculated the d1/d2 ratio. At least 30 adults were examined in each experiment. Imaginal disc immunostaining
30 Imaginal discs from third instar wandering larvae were dissected in PBS, fixed by incubation for 30 min at room temperature (RT) in 4% paraformaldehyde, washed and permeabilized by three incubations for 10 minutes each in PBS + 0.1% Tween (PBST). They were blocked by incubation for one hour in PBST + 2% BSA, then overnight at 4 C with the primary antibody. They were washed three times, for 10 minutes each with PBST, blocked by incubation for 1 hour in PBST + 2% BSA and incubated for 3 hours at RT with the secondary antibody (in PBST + 2% BSA). Finally, they were rinsed three times, for 10 minutes each, in PBST and mounted in Citifluor. Confocal imaging Cell culture images were acquired with a Yokogawa spinning disk confocal head coupled to a Leica inverted microscope (Leica, DMIRB). Excitation was achieved with an Argon-Krypton 75 mw laser (Melles Griot). Images were acquired with a CoolSnap HQ camera (Princeton Instruments). The whole set-up was driven by Metamorph 6.3 (Universal Imaging). The integration and wavelength switch module was designed by the firm Errol and the Institut Jacques Monod imaging facility. Supplemental bibliographie 1. Brand, A.H., Manoukian, A.S., and Perrimon, N. (1994). Ectopic expression in Drosophila. Methods Cell Biol 44, Rudolph, M.J., Amodeo, G.A., Bai, Y., and Tong, L. (2005). Crystal structure of the protein kinase domain of yeast AMP-activated protein kinase Snf1. Biochem Biophys Res Commun 337, Ascano, M., Jr., Nybakken, K.E., Sosinski, J., Stegman, M.A., and Robbins, D.J. (2002). The carboxylterminal domain of the protein kinase fused can function as a dominant inhibitor of hedgehog signaling. Mol Cell Biol 22, Ascano, M., Jr., and Robbins, D.J. (2004). An intramolecular association between two domains of the protein kinase Fused is necessary for Hedgehog signaling. Mol Cell Biol 24, Monnier, V., Dussillol, F., Alves, G., Lamour, I.C., and Plessis, A. (1998). Suppressor of fused links fused and Cubitus interruptus on the hedgehog signalling pathway. Curr Biol 8, Pham, A., Therond, P., Alves, G., Tournier, F.B., Busson, D., Lamour Isnard, C., Bouchon, B.L., Préat, T., and Tricoire, H. (1995). The Suppressor of fused gene encodes a novel PEST protein involved in Drosophila segment polarity establishment. Genetics 140, Ho, K.S., Suyama, K., Fish, M., and Scott, M.P. (2005). Differential regulation of Hedgehog target gene transcription by Costal2 and Suppressor of Fused. Development 132,
31 8. Dussillol-Godar, F., Brissard-Zahraoui, J., Limbourg-Bouchon, B., Boucher, D., Fouix, S., Lamour- Isnard, C., Plessis, A., and Busson, D. (2006). Modulation of the Suppressor of fused protein regulates the Hedgehog signaling pathway in Drosophila embryo and imaginal discs. Dev Biol. 9. Malpel, S., Claret, S., Sanial, M., Brigui, A., Piolot, T., Daviet, L., Martin-Lanneree, S., and Plessis, A. (2007). The last 59 amino acids of Smoothened cytoplasmic tail directly bind the protein kinase Fused and negatively regulate the Hedgehog pathway. Dev Biol 303, Zhu, A.J., Zheng, L., Suyama, K., and Scott, M.P. (2003). Altered localization of Drosophila Smoothened protein activates Hedgehog signal transduction. Genes Dev 17, Taipale, J., Cooper, M.K., Maiti, T., and Beachy, P.A. (2002). Patched acts catalytically to suppress the activity of Smoothened. Nature 418, Fukumoto, T., Watanabe-Fukunaga, R., Fujisawa, K., Nagata, S., and Fukunaga, R. (2001). The fused protein kinase regulates Hedgehog-stimulated transcriptional activation in Drosophila Schneider 2 cells. J Biol Chem 276, Lee, T., and Luo, L. (2001). Mosaic analysis with a repressible cell marker (MARCM) for Drosophila neural development. Trends Neurosci 24, Capdevila, J., and Guerrero, I. (1994). Targeted expression of the signaling molecule decapentaplegic induces pattern duplications and growth alterations in Drosophila wings. Embo J 13, Brand, A.H., and Perrimon, N. (1993). Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118, Pignoni, F., and Zipursky, S.L. (1997). Induction of Drosophila eye development by decapentaplegic. Development 124, Therond, P.P., Knight, J.D., Kornberg, T.B., and Bishop, J.M. (1996). Phosphorylation of the fused protein kinase in response to signaling from hedgehog. Proc Natl Acad Sci U S A 93, van den Heuvel, M., and Ingham, P.W. (1996). smoothened encodes a receptor-like serpentin protein required for hedgehog signalling. Nature 382,
32 Supplemental figure 1
33 Supplemental figure 2
34 Supplemental figure 3
35 Supplemental figure 4
36 Supplemental figure 5
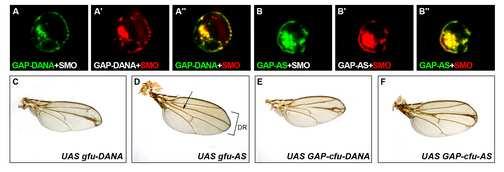
37 Supplemental figure 6
38 Supplemental figure 7
39 Supplemental figure 8
40 Supplemental figure 9
The last 59 amino acids of Smoothened cytoplasmic tail directly bind the protein kinase Fused and negatively regulate the Hedgehog pathway
 Developmental Biology 303 (2007) 121 133 www.elsevier.com/locate/ydbio The last 59 amino acids of Smoothened cytoplasmic tail directly bind the protein kinase Fused and negatively regulate the Hedgehog
Developmental Biology 303 (2007) 121 133 www.elsevier.com/locate/ydbio The last 59 amino acids of Smoothened cytoplasmic tail directly bind the protein kinase Fused and negatively regulate the Hedgehog
THE Hedgehog (Hh) signaling pathway regulates
 signaling pathway regulates") Copyright Ó 2006 by the Genetics Society of America DOI: 10.1534/genetics.106.061036 The Contributions of Protein Kinase A and Smoothened Phosphorylation to Hedgehog Signal Transduction in Drosophila melanogaster
Copyright Ó 2006 by the Genetics Society of America DOI: 10.1534/genetics.106.061036 The Contributions of Protein Kinase A and Smoothened Phosphorylation to Hedgehog Signal Transduction in Drosophila melanogaster
Segment boundary formation in Drosophila embryos
 Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
Segment boundary formation in Drosophila embryos Development 130, August 2003 Camilla W. Larsen, Elizabeth Hirst, Cyrille Alexandre and Jean Paul Vincent 1. Introduction: - Segment boundary formation:
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
Research article. Summary The sex determination master switch, Sex-lethal (Sxl), Introduction
, Introduction") 6101 A positive role for Patched in Hedgehog signaling revealed by the intracellular trafficking of Sex-lethal, the Drosophila sex determination master switch Jamila I. Horabin 1, *, Sabrina Walthall 1,
6101 A positive role for Patched in Hedgehog signaling revealed by the intracellular trafficking of Sex-lethal, the Drosophila sex determination master switch Jamila I. Horabin 1, *, Sabrina Walthall 1,
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
4) Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.
 Please cite Dagda et al J Biol Chem 284: , for any publications or presentations resulting from use or modification of the macro.") Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Supplement Figure S1. Algorithmic quantification of mitochondrial morphology in SH- SY5Y cells treated with known fission/fusion mediators. Parental SH-SY5Y cells were transiently transfected with an empty
Report. Costal2 Functions as a Kinesin-like Protein in the Hedgehog Signal Transduction Pathway
 Current Biology 18, 1215 1220, August 26, 2008 ª2008 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2008.07.026 Costal2 Functions as a Kinesin-like Protein in the Hedgehog Signal Transduction Pathway
Current Biology 18, 1215 1220, August 26, 2008 ª2008 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2008.07.026 Costal2 Functions as a Kinesin-like Protein in the Hedgehog Signal Transduction Pathway
A large complex containing Patched and Smoothened initiates Hedgehog signaling in Drosophila
 826 Research Article A large complex containing Patched and Smoothened initiates Hedgehog signaling in Drosophila Sabrina L. Walthall 1, Michelle Moses 1 and Jamila I. Horabin 2, * 1 Department of Biochemistry
826 Research Article A large complex containing Patched and Smoothened initiates Hedgehog signaling in Drosophila Sabrina L. Walthall 1, Michelle Moses 1 and Jamila I. Horabin 2, * 1 Department of Biochemistry
Supplementary Information
 Supplementary Information Supplementary Figure 1. JAK/STAT in early wing development (a-f) Wing primordia of second instar larvae of the indicated genotypes labeled to visualize expression of upd mrna
Supplementary Information Supplementary Figure 1. JAK/STAT in early wing development (a-f) Wing primordia of second instar larvae of the indicated genotypes labeled to visualize expression of upd mrna
The function of engrailed and the specification of Drosophila wing pattern
 Development 121, 3447-3456 (1995) Printed in Great Britain The Company of Biologists Limited 1995 3447 The function of engrailed and the specification of Drosophila wing pattern Isabel Guillén 1, *, Jose
Development 121, 3447-3456 (1995) Printed in Great Britain The Company of Biologists Limited 1995 3447 The function of engrailed and the specification of Drosophila wing pattern Isabel Guillén 1, *, Jose
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
DOI: 10.1038/ncb3267 Supplementary Figure 1 A group of genes required for formation or orientation of annular F-actin bundles and aecm ridges: RNAi phenotypes and their validation by standard mutations.
Midterm 1. Average score: 74.4 Median score: 77
 Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Midterm 1 Average score: 74.4 Median score: 77 NAME: TA (circle one) Jody Westbrook or Jessica Piel Section (circle one) Tue Wed Thur MCB 141 First Midterm Feb. 21, 2008 Only answer 4 of these 5 problems.
Axis determination in flies. Sem 9.3.B.5 Animal Science
 Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Drosophila. Cubitus interruptus-independent transduction of the Hedgehog signal in
 Development 127, 5509-5522 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV7818 5509 Cubitus interruptus-independent transduction of the Hedgehog signal in Drosophila Armel Gallet
Development 127, 5509-5522 (2000) Printed in Great Britain The Company of Biologists Limited 2000 DEV7818 5509 Cubitus interruptus-independent transduction of the Hedgehog signal in Drosophila Armel Gallet
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Nature Neuroscience: doi: /nn.2662
 Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
targets. clustering show that different complex pathway
 Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Supplementary Figure 1. CLICR allows clustering and activation of cytoplasmic protein targets. (a, b) Upon light activation, the Cry2 (red) and LRP6c (green) components co-cluster due to the heterodimeric
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
SUPPLEMENTARY INFORMATION
 GP2 Type I-piliated bacteria FAE M cell M cell pocket idc T cell mdc Generation of antigenspecific T cells Induction of antigen-specific mucosal immune response Supplementary Figure 1 Schematic diagram
GP2 Type I-piliated bacteria FAE M cell M cell pocket idc T cell mdc Generation of antigenspecific T cells Induction of antigen-specific mucosal immune response Supplementary Figure 1 Schematic diagram
Drosophila Life Cycle
 Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Sonic hedgehog (Shh) signalling in the rabbit embryo
 signalling in the rabbit embryo") Sonic hedgehog (Shh) signalling in the rabbit embryo In the first part of this thesis work the physical properties of cilia-driven leftward flow were characterised in the rabbit embryo. Since its discovery
Sonic hedgehog (Shh) signalling in the rabbit embryo In the first part of this thesis work the physical properties of cilia-driven leftward flow were characterised in the rabbit embryo. Since its discovery
SUPPLEMENTARY INFORMATION
 med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
med!1,2 Wild-type (N2) end!3 elt!2 5 1 15 Time (minutes) 5 1 15 Time (minutes) med!1,2 end!3 5 1 15 Time (minutes) elt!2 5 1 15 Time (minutes) Supplementary Figure 1: Number of med-1,2, end-3, end-1 and
Smart Bolometer: Toward Monolithic Bolometer with Smart Functions
 Smart Bolometer: Toward Monolithic Bolometer with Smart Functions Matthieu Denoual, Gilles Allègre, Patrick Attia, Olivier De Sagazan To cite this version: Matthieu Denoual, Gilles Allègre, Patrick Attia,
Smart Bolometer: Toward Monolithic Bolometer with Smart Functions Matthieu Denoual, Gilles Allègre, Patrick Attia, Olivier De Sagazan To cite this version: Matthieu Denoual, Gilles Allègre, Patrick Attia,
Supporting Online Material for
 www.sciencemag.org/cgi/content/full/1121356/dc1 Supporting Online Material for Polar PIN Localization Directs Auxin Flow in Plants Justyna Wiśniewska, Jian Xu, Daniela Seifertová, Philip B. Brewer, Kamil
www.sciencemag.org/cgi/content/full/1121356/dc1 Supporting Online Material for Polar PIN Localization Directs Auxin Flow in Plants Justyna Wiśniewska, Jian Xu, Daniela Seifertová, Philip B. Brewer, Kamil
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
IMPROVEMENTS OF THE VARIABLE THERMAL RESISTANCE
 IMPROVEMENTS OF THE VARIABLE THERMAL RESISTANCE V. Szekely, S. Torok, E. Kollar To cite this version: V. Szekely, S. Torok, E. Kollar. IMPROVEMENTS OF THE VARIABLE THERMAL RESIS- TANCE. THERMINIC 2007,
IMPROVEMENTS OF THE VARIABLE THERMAL RESISTANCE V. Szekely, S. Torok, E. Kollar To cite this version: V. Szekely, S. Torok, E. Kollar. IMPROVEMENTS OF THE VARIABLE THERMAL RESIS- TANCE. THERMINIC 2007,
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila
 Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
Baz, Par-6 and apkc are not required for axon or dendrite specification in Drosophila Melissa M. Rolls and Chris Q. Doe, Inst. Neurosci and Inst. Mol. Biol., HHMI, Univ. Oregon, Eugene, Oregon 97403 Correspondence
Why Flies? stages of embryogenesis. The Fly in History
 The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
The Fly in History 1859 Darwin 1866 Mendel c. 1890 Driesch, Roux (experimental embryology) 1900 rediscovery of Mendel (birth of genetics) 1910 first mutant (white) (Morgan) 1913 first genetic map (Sturtevant
AP Biology Gene Regulation and Development Review
 AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
Supplementary Materials for
 www.sciencemag.org/cgi/content/full/science.1244624/dc1 Supplementary Materials for Cytoneme-Mediated Contact-Dependent Transport of the Drosophila Decapentaplegic Signaling Protein Sougata Roy, Hai Huang,
www.sciencemag.org/cgi/content/full/science.1244624/dc1 Supplementary Materials for Cytoneme-Mediated Contact-Dependent Transport of the Drosophila Decapentaplegic Signaling Protein Sougata Roy, Hai Huang,
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Establishing positional information through gradient dynamics
 Extra view Fly 4:4, 273-277; October/November/December 2010; 2010 Landes Bioscience Extra view Establishing positional information through gradient dynamics A lesson from the Hedgehog signaling pathway
Extra view Fly 4:4, 273-277; October/November/December 2010; 2010 Landes Bioscience Extra view Establishing positional information through gradient dynamics A lesson from the Hedgehog signaling pathway
Supplementary Figure 1
 Supplementary Figure 1 Supplementary Figure 1. HSP21 expression in 35S:HSP21 and hsp21 knockdown plants. (a) Since no T- DNA insertion line for HSP21 is available in the publicly available T-DNA collections,
Supplementary Figure 1 Supplementary Figure 1. HSP21 expression in 35S:HSP21 and hsp21 knockdown plants. (a) Since no T- DNA insertion line for HSP21 is available in the publicly available T-DNA collections,
Supplemental Data. Chen and Thelen (2010). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Data. Chen and Thelen (2010). Plant Cell 10.1105/tpc.109.071837 1 C Total 5 kg 20 kg 100 kg Transmission Image 100 kg soluble pdtpi-gfp Plastid (PDH-alpha) Mito (PDH-alpha) GFP Image vector
Supplemental Data. Chen and Thelen (2010). Plant Cell 10.1105/tpc.109.071837 1 C Total 5 kg 20 kg 100 kg Transmission Image 100 kg soluble pdtpi-gfp Plastid (PDH-alpha) Mito (PDH-alpha) GFP Image vector
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins
 13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
13-3. Synthesis-Secretory pathway: Sort lumenal proteins, Secrete proteins, Sort membrane proteins Molecular sorting: specific budding, vesicular transport, fusion 1. Why is this important? A. Form and
Lipniacki 2004 Ground Truth
 Abstract Lipniacki 2004 Ground Truth The two-feedback-loop regulatory module of nuclear factor kb (NF-kB) signaling pathway is modeled by means of ordinary differential equations. signaling pathway: https://en.wikipedia.org/wiki/signaling_pathway
Abstract Lipniacki 2004 Ground Truth The two-feedback-loop regulatory module of nuclear factor kb (NF-kB) signaling pathway is modeled by means of ordinary differential equations. signaling pathway: https://en.wikipedia.org/wiki/signaling_pathway
Determination of absorption characteristic of materials on basis of sound intensity measurement
 Determination of absorption characteristic of materials on basis of sound intensity measurement R. Prascevic, A. Milosevic, S. Cvetkovic To cite this version: R. Prascevic, A. Milosevic, S. Cvetkovic.
Determination of absorption characteristic of materials on basis of sound intensity measurement R. Prascevic, A. Milosevic, S. Cvetkovic To cite this version: R. Prascevic, A. Milosevic, S. Cvetkovic.
Nature Biotechnology: doi: /nbt Supplementary Figure 1. Overexpression of YFP::GPR-1 in the germline.
 Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
A new simple recursive algorithm for finding prime numbers using Rosser s theorem
 A new simple recursive algorithm for finding prime numbers using Rosser s theorem Rédoane Daoudi To cite this version: Rédoane Daoudi. A new simple recursive algorithm for finding prime numbers using Rosser
A new simple recursive algorithm for finding prime numbers using Rosser s theorem Rédoane Daoudi To cite this version: Rédoane Daoudi. A new simple recursive algorithm for finding prime numbers using Rosser
Easter bracelets for years
 Easter bracelets for 5700000 years Denis Roegel To cite this version: Denis Roegel. Easter bracelets for 5700000 years. [Research Report] 2014. HAL Id: hal-01009457 https://hal.inria.fr/hal-01009457
Easter bracelets for 5700000 years Denis Roegel To cite this version: Denis Roegel. Easter bracelets for 5700000 years. [Research Report] 2014. HAL Id: hal-01009457 https://hal.inria.fr/hal-01009457
7.06 Cell Biology EXAM #3 April 21, 2005
 7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
DEM modeling of penetration test in static and dynamic conditions
 DEM modeling of penetration test in static and dynamic conditions Quoc Anh Tran, Bastien Chevalier, Pierre Breul To cite this version: Quoc Anh Tran, Bastien Chevalier, Pierre Breul. DEM modeling of penetration
DEM modeling of penetration test in static and dynamic conditions Quoc Anh Tran, Bastien Chevalier, Pierre Breul To cite this version: Quoc Anh Tran, Bastien Chevalier, Pierre Breul. DEM modeling of penetration
SUPPLEMENTARY INFORMATION
 Supplementary Figure 1 Sns and Duf co-localise in embryonic nephrocytes a-c, Wild-type stage 11 (a),14 (b) and 16 (c) embryos stained with anti-duf (green) and anti-sns (red). Higher magnification images
Supplementary Figure 1 Sns and Duf co-localise in embryonic nephrocytes a-c, Wild-type stage 11 (a),14 (b) and 16 (c) embryos stained with anti-duf (green) and anti-sns (red). Higher magnification images
Segmenting the fly embryo: logical analysis of the role of the Segment Polarity cross-regulatory module
 Int. J. Dev. Biol. 52: 1059-1075 (2008) doi: 10.1387/ijdb.072439ls THE INTERNATIONAL JOURNAL OF DEVELOPMENTAL BIOLOGY www.intjdevbiol.com Segmenting the fly embryo: logical analysis of the role of the
Int. J. Dev. Biol. 52: 1059-1075 (2008) doi: 10.1387/ijdb.072439ls THE INTERNATIONAL JOURNAL OF DEVELOPMENTAL BIOLOGY www.intjdevbiol.com Segmenting the fly embryo: logical analysis of the role of the
Can we reduce health inequalities? An analysis of the English strategy ( )
") Can we reduce health inequalities? An analysis of the English strategy (1997-2010) Johan P Mackenbach To cite this version: Johan P Mackenbach. Can we reduce health inequalities? An analysis of the English
Can we reduce health inequalities? An analysis of the English strategy (1997-2010) Johan P Mackenbach To cite this version: Johan P Mackenbach. Can we reduce health inequalities? An analysis of the English
downstream (0.8 kb) homologous sequences to the genomic locus of DIC. A DIC mutant strain (ro- 6
 homologous sequences to the genomic locus of DIC. A DIC mutant strain (ro- 6") A B C D ts Figure S1 Generation of DIC- mcherry expressing N.crassa strain. A. N. crassa colony morphology. When a cot1 (top, left panel) strain is grown at permissive temperature (25 C), it exhibits straight
A B C D ts Figure S1 Generation of DIC- mcherry expressing N.crassa strain. A. N. crassa colony morphology. When a cot1 (top, left panel) strain is grown at permissive temperature (25 C), it exhibits straight
Methylation-associated PHOX2B gene silencing is a rare event in human neuroblastoma.
 Methylation-associated PHOX2B gene silencing is a rare event in human neuroblastoma. Loïc De Pontual, Delphine Trochet, Franck Bourdeaut, Sophie Thomas, Heather Etchevers, Agnes Chompret, Véronique Minard,
Methylation-associated PHOX2B gene silencing is a rare event in human neuroblastoma. Loïc De Pontual, Delphine Trochet, Franck Bourdeaut, Sophie Thomas, Heather Etchevers, Agnes Chompret, Véronique Minard,
Full-order observers for linear systems with unknown inputs
 Full-order observers for linear systems with unknown inputs Mohamed Darouach, Michel Zasadzinski, Shi Jie Xu To cite this version: Mohamed Darouach, Michel Zasadzinski, Shi Jie Xu. Full-order observers
Full-order observers for linear systems with unknown inputs Mohamed Darouach, Michel Zasadzinski, Shi Jie Xu To cite this version: Mohamed Darouach, Michel Zasadzinski, Shi Jie Xu. Full-order observers
Characterization of the local Electrical Properties of Electrical Machine Parts with non-trivial Geometry
 Characterization of the local Electrical Properties of Electrical Machine Parts with non-trivial Geometry Laure Arbenz, Abdelkader Benabou, Stéphane Clenet, Jean Claude Mipo, Pierre Faverolle To cite this
Characterization of the local Electrical Properties of Electrical Machine Parts with non-trivial Geometry Laure Arbenz, Abdelkader Benabou, Stéphane Clenet, Jean Claude Mipo, Pierre Faverolle To cite this
In order to confirm the contribution of diffusion to the FRAP recovery curves of
 Fonseca et al., Supplementary Information FRAP data analysis ) Contribution of Diffusion to the recovery curves In order to confirm the contribution of diffusion to the FRAP recovery curves of PH::GFP
Fonseca et al., Supplementary Information FRAP data analysis ) Contribution of Diffusion to the recovery curves In order to confirm the contribution of diffusion to the FRAP recovery curves of PH::GFP
Developmental genetics: finding the genes that regulate development
 Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Developmental Biology BY1101 P. Murphy Lecture 9 Developmental genetics: finding the genes that regulate development Introduction The application of genetic analysis and DNA technology to the study of
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
Studying the effect of cell division on expression patterns of the segment polarity genes
 Studying the effect of cell division on expression patterns of the segment polarity genes Madalena Chaves and Réka Albert Abstract The segment polarity gene family, and its gene regulatory network, is
Studying the effect of cell division on expression patterns of the segment polarity genes Madalena Chaves and Réka Albert Abstract The segment polarity gene family, and its gene regulatory network, is
Supplementary Figure 1. Markedly decreased numbers of marginal zone B cells in DOCK8 mutant mice Supplementary Figure 2.
 Supplementary Figure 1. Markedly decreased numbers of marginal zone B cells in DOCK8 mutant mice. Percentage of marginal zone B cells in the spleen of wild-type mice (+/+), mice homozygous for cpm or pri
Supplementary Figure 1. Markedly decreased numbers of marginal zone B cells in DOCK8 mutant mice. Percentage of marginal zone B cells in the spleen of wild-type mice (+/+), mice homozygous for cpm or pri
Dpp Signaling Activity Requires Pentagone to Scale with Tissue Size in the Growing Drosophila Wing Imaginal Disc
 Dpp Signaling Activity Requires Pentagone to Scale with Tissue Size in the Growing Drosophila Wing Imaginal Disc Fisun Hamaratoglu 1. *, Aitana Morton de Lachapelle 2,3., George Pyrowolakis 4,5, Sven Bergmann
Dpp Signaling Activity Requires Pentagone to Scale with Tissue Size in the Growing Drosophila Wing Imaginal Disc Fisun Hamaratoglu 1. *, Aitana Morton de Lachapelle 2,3., George Pyrowolakis 4,5, Sven Bergmann
Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering
 Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering by Valverde et. Al Published in Science 2004 Presented by Boyana Grigorova CBMG 688R Feb. 12, 2007 Circadian Rhythms: The Clock Within
Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering by Valverde et. Al Published in Science 2004 Presented by Boyana Grigorova CBMG 688R Feb. 12, 2007 Circadian Rhythms: The Clock Within
Specificities of Dishevelled
 Casein Kinase Iε Modulates the Signaling Specificities of Dishevelled Feng Cong, Liang Schweizer and Harold Varmus Mol. Cell. Biol. 2004, 24(5):2000. DOI: 10.1128/MCB.24.5.2000-2011.2004. REFERENCES CONTENT
Casein Kinase Iε Modulates the Signaling Specificities of Dishevelled Feng Cong, Liang Schweizer and Harold Varmus Mol. Cell. Biol. 2004, 24(5):2000. DOI: 10.1128/MCB.24.5.2000-2011.2004. REFERENCES CONTENT
Types of biological networks. I. Intra-cellurar networks
 Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Types of biological networks I. Intra-cellurar networks 1 Some intra-cellular networks: 1. Metabolic networks 2. Transcriptional regulation networks 3. Cell signalling networks 4. Protein-protein interaction
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
A note on the computation of the fraction of smallest denominator in between two irreducible fractions
 A note on the computation of the fraction of smallest denominator in between two irreducible fractions Isabelle Sivignon To cite this version: Isabelle Sivignon. A note on the computation of the fraction
A note on the computation of the fraction of smallest denominator in between two irreducible fractions Isabelle Sivignon To cite this version: Isabelle Sivignon. A note on the computation of the fraction
 DOI: 10.1038/ncb2819 Gαi3 / Actin / Acetylated Tubulin Gαi3 / Actin / Acetylated Tubulin a a Gαi3 a Actin Gαi3 WT Gαi3 WT Gαi3 WT b b Gαi3 b Actin Gαi3 KO Gαi3 KO Gαi3 KO # # Figure S1 Loss of protein
DOI: 10.1038/ncb2819 Gαi3 / Actin / Acetylated Tubulin Gαi3 / Actin / Acetylated Tubulin a a Gαi3 a Actin Gαi3 WT Gαi3 WT Gαi3 WT b b Gαi3 b Actin Gαi3 KO Gαi3 KO Gαi3 KO # # Figure S1 Loss of protein
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
Basins of Attraction Plasticity of a Strange Attractor with a Swirling Scroll
 Basins of Attraction Plasticity of a Strange Attractor with a Swirling Scroll Safieddine Bouali To cite this version: Safieddine Bouali. Basins of Attraction Plasticity of a Strange Attractor with a Swirling
Basins of Attraction Plasticity of a Strange Attractor with a Swirling Scroll Safieddine Bouali To cite this version: Safieddine Bouali. Basins of Attraction Plasticity of a Strange Attractor with a Swirling
Supplemental Data. Perrella et al. (2013). Plant Cell /tpc
. Plant Cell /tpc") Intensity Intensity Intensity Intensity Intensity Intensity 150 50 150 0 10 20 50 C 150 0 10 20 50 D 0 10 20 Distance (μm) 50 20 40 E 50 F 0 10 20 50 0 15 30 Distance (μm) Supplemental Figure 1: Co-localization
Intensity Intensity Intensity Intensity Intensity Intensity 150 50 150 0 10 20 50 C 150 0 10 20 50 D 0 10 20 Distance (μm) 50 20 40 E 50 F 0 10 20 50 0 15 30 Distance (μm) Supplemental Figure 1: Co-localization
Thomas Lugand. To cite this version: HAL Id: tel
 Contribution à la Modélisation et à l Optimisation de la Machine Asynchrone Double Alimentation pour des Applications Hydrauliques de Pompage Turbinage Thomas Lugand To cite this version: Thomas Lugand.
Contribution à la Modélisation et à l Optimisation de la Machine Asynchrone Double Alimentation pour des Applications Hydrauliques de Pompage Turbinage Thomas Lugand To cite this version: Thomas Lugand.
Comment on: Sadi Carnot on Carnot s theorem.
 Comment on: Sadi Carnot on Carnot s theorem. Jacques Arnaud, Laurent Chusseau, Fabrice Philippe To cite this version: Jacques Arnaud, Laurent Chusseau, Fabrice Philippe. Comment on: Sadi Carnot on Carnot
Comment on: Sadi Carnot on Carnot s theorem. Jacques Arnaud, Laurent Chusseau, Fabrice Philippe To cite this version: Jacques Arnaud, Laurent Chusseau, Fabrice Philippe. Comment on: Sadi Carnot on Carnot
Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway
 Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway Justin Hogan, Meagan Valentine, Chris Cox, Kristy Doyle, Simon Collier* Department
Two Frizzled Planar Cell Polarity Signals in the Drosophila Wing Are Differentially Organized by the Fat/Dachsous Pathway Justin Hogan, Meagan Valentine, Chris Cox, Kristy Doyle, Simon Collier* Department
Interactions of an eddy current sensor and a multilayered structure
 Interactions of an eddy current sensor and a multilayered structure Thanh Long Cung, Pierre-Yves Joubert, Eric Vourc H, Pascal Larzabal To cite this version: Thanh Long Cung, Pierre-Yves Joubert, Eric
Interactions of an eddy current sensor and a multilayered structure Thanh Long Cung, Pierre-Yves Joubert, Eric Vourc H, Pascal Larzabal To cite this version: Thanh Long Cung, Pierre-Yves Joubert, Eric
Stochastic simulations
 Stochastic simulations Application to molecular networks Literature overview Noise in genetic networks Origins How to measure and distinguish between the two types of noise (intrinsic vs extrinsic)? What
Stochastic simulations Application to molecular networks Literature overview Noise in genetic networks Origins How to measure and distinguish between the two types of noise (intrinsic vs extrinsic)? What
Pattern formation: Wingless on the move Robert Howes and Sarah Bray
 R222 Dispatch Pattern formation: Wingless on the move Robert Howes and Sarah Bray Wingless is a key morphogen in Drosophila. Although it is evident that Wingless acts at a distance from its site of synthesis,
R222 Dispatch Pattern formation: Wingless on the move Robert Howes and Sarah Bray Wingless is a key morphogen in Drosophila. Although it is evident that Wingless acts at a distance from its site of synthesis,
Principles of Experimental Embryology
 Biology 4361 Developmental Biology Principles of Experimental Embryology September 19, 2006 Major Research Questions How do forces outside the embryo affect its development? (Environmental Developmental
Biology 4361 Developmental Biology Principles of Experimental Embryology September 19, 2006 Major Research Questions How do forces outside the embryo affect its development? (Environmental Developmental
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its
 The Wnt/ -catenin signalling pathway and its") Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Zool 3200: Cell Biology Exam 5 4/27/15
 Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
SUPPLEMENTARY INFORMATION
 DOI: 10.1038/ncb2647 Figure S1 Other Rab GTPases do not co-localize with the ER. a, Cos-7 cells cotransfected with an ER luminal marker (either KDEL-venus or mch-kdel) and mch-tagged human Rab5 (mch-rab5,
DOI: 10.1038/ncb2647 Figure S1 Other Rab GTPases do not co-localize with the ER. a, Cos-7 cells cotransfected with an ER luminal marker (either KDEL-venus or mch-kdel) and mch-tagged human Rab5 (mch-rab5,
Supplementary Figures
 Supplementary Figures Supplementary Figure 1. Purification of yeast CKM. (a) Silver-stained SDS-PAGE analysis of CKM purified through a TAP-tag engineered into the Cdk8 C-terminus. (b) Kinase activity
Supplementary Figures Supplementary Figure 1. Purification of yeast CKM. (a) Silver-stained SDS-PAGE analysis of CKM purified through a TAP-tag engineered into the Cdk8 C-terminus. (b) Kinase activity
Supplemental Information. The Mitochondrial Fission Receptor MiD51. Requires ADP as a Cofactor
 Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
The majority of cells in the nervous system arise during the embryonic and early post
 Introduction Introduction The majority of cells in the nervous system arise during the embryonic and early post natal period. These cells are derived from population of neural stem cells first shown by
Introduction Introduction The majority of cells in the nervous system arise during the embryonic and early post natal period. These cells are derived from population of neural stem cells first shown by
Case report on the article Water nanoelectrolysis: A simple model, Journal of Applied Physics (2017) 122,
 122,") Case report on the article Water nanoelectrolysis: A simple model, Journal of Applied Physics (2017) 122, 244902 Juan Olives, Zoubida Hammadi, Roger Morin, Laurent Lapena To cite this version: Juan Olives,
Case report on the article Water nanoelectrolysis: A simple model, Journal of Applied Physics (2017) 122, 244902 Juan Olives, Zoubida Hammadi, Roger Morin, Laurent Lapena To cite this version: Juan Olives,
Lecture 10: Cyclins, cyclin kinases and cell division
 Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
MCB 141 Midterm I Feb. 14, 2012
 Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 14, 2012 Question #1 Question #2 Question #3 Question #4 BONUS / 28 pts / 27 pts / 25 pts / 20 pts / 1 pt TOTAL / 100 pts
Write your name and student ID# on EVERY PAGE of your exam MCB 141 Midterm I Feb. 14, 2012 Question #1 Question #2 Question #3 Question #4 BONUS / 28 pts / 27 pts / 25 pts / 20 pts / 1 pt TOTAL / 100 pts
Morphogens in biological development: Drosophila example
 LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
LSM5194 Morphogens in biological development: Drosophila example Lecture 29 The concept of morphogen gradients The concept of morphogens was proposed by L. Wolpert as a part of the positional information
Cell Cell Communication in Development
 Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Role of Mitochondrial Remodeling in Programmed Cell Death in
 Developmental Cell, Vol. 12 Supplementary Data Role of Mitochondrial Remodeling in Programmed Cell Death in Drosophila melanogaster Gaurav Goyal, Brennan Fell, Apurva Sarin, Richard J. Youle, V. Sriram.
Developmental Cell, Vol. 12 Supplementary Data Role of Mitochondrial Remodeling in Programmed Cell Death in Drosophila melanogaster Gaurav Goyal, Brennan Fell, Apurva Sarin, Richard J. Youle, V. Sriram.
The Hedgehog Response Network: Sensors, Switches, and Routers. Lawrence Lum and Philip A. Beachy*
 The Hedgehog Response Network: Sensors, Switches, and Routers Lawrence Lum and Philip. Beachy* The Hedgehog () signaling pathway is intimately linked to cell growth and differentiation, with normal roles
The Hedgehog Response Network: Sensors, Switches, and Routers Lawrence Lum and Philip. Beachy* The Hedgehog () signaling pathway is intimately linked to cell growth and differentiation, with normal roles
Supplementary Figure 1. Nature Genetics: doi: /ng.3848
 Supplementary Figure 1 Phenotypes and epigenetic properties of Fab2L flies. A- Phenotypic classification based on eye pigment levels in Fab2L male (orange bars) and female (yellow bars) flies (n>150).
Supplementary Figure 1 Phenotypes and epigenetic properties of Fab2L flies. A- Phenotypic classification based on eye pigment levels in Fab2L male (orange bars) and female (yellow bars) flies (n>150).
Richik N. Ghosh, Linnette Grove, and Oleg Lapets ASSAY and Drug Development Technologies 2004, 2:
 1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
1 3/1/2005 A Quantitative Cell-Based High-Content Screening Assay for the Epidermal Growth Factor Receptor-Specific Activation of Mitogen-Activated Protein Kinase Richik N. Ghosh, Linnette Grove, and Oleg
Early detection of thermal contrast in pulsed stimulated thermography
 Early detection of thermal contrast in pulsed stimulated thermography J.-C. Krapez, F. Lepoutre, D. Balageas To cite this version: J.-C. Krapez, F. Lepoutre, D. Balageas. Early detection of thermal contrast
Early detection of thermal contrast in pulsed stimulated thermography J.-C. Krapez, F. Lepoutre, D. Balageas To cite this version: J.-C. Krapez, F. Lepoutre, D. Balageas. Early detection of thermal contrast
Under the Radar Screen: How Bugs Trick Our Immune Defenses
 Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 2: Phagocytosis Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research Salmonella Gram negative bacteria
Under the Radar Screen: How Bugs Trick Our Immune Defenses Session 2: Phagocytosis Marie-Eve Paquet and Gijsbert Grotenbreg Whitehead Institute for Biomedical Research Salmonella Gram negative bacteria
Biol403 - Receptor Serine/Threonine Kinases
 Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Biol403 - Receptor Serine/Threonine Kinases The TGFβ (transforming growth factorβ) family of growth factors TGFβ1 was first identified as a transforming factor; however, it is a member of a family of structurally
Enhanced zinc-finger-nuclease activity with improved obligate heterodimeric architectures
 Nature Methods Enhanced zinc-finger-nuclease activity with improved obligate heterodimeric architectures Yannick Doyon, Thuy D Vo, Matthew C Mendel, Shon G Greenberg, Jianbin Wang, Danny F Xia, Jeffrey
Nature Methods Enhanced zinc-finger-nuclease activity with improved obligate heterodimeric architectures Yannick Doyon, Thuy D Vo, Matthew C Mendel, Shon G Greenberg, Jianbin Wang, Danny F Xia, Jeffrey
Natural convection of magnetic fluid inside a cubical enclosure under magnetic gravity compensation
 Natural convection of magnetic fluid inside a cubical enclosure under magnetic gravity compensation Zuo-Sheng Lei, Sheng-Yang Song, Chun-Long Xu, Jia-Hong Guo To cite this version: Zuo-Sheng Lei, Sheng-Yang
Natural convection of magnetic fluid inside a cubical enclosure under magnetic gravity compensation Zuo-Sheng Lei, Sheng-Yang Song, Chun-Long Xu, Jia-Hong Guo To cite this version: Zuo-Sheng Lei, Sheng-Yang
Water Vapour Effects in Mass Measurement
 Water Vapour Effects in Mass Measurement N.-E. Khélifa To cite this version: N.-E. Khélifa. Water Vapour Effects in Mass Measurement. Measurement. Water Vapour Effects in Mass Measurement, May 2007, Smolenice,
Water Vapour Effects in Mass Measurement N.-E. Khélifa To cite this version: N.-E. Khélifa. Water Vapour Effects in Mass Measurement. Measurement. Water Vapour Effects in Mass Measurement, May 2007, Smolenice,
Heather Currinn, Benjamin Guscott, Zita Balklava, Alice Rothnie and Thomas Wassmer*
 Online Resources APP controls the formation of PI(3,5)P 2 vesicles through its binding of the PIKfyve complex. Cellular and Molecular Life Sciences Heather Currinn, Benjamin Guscott, Zita Balklava, Alice
Online Resources APP controls the formation of PI(3,5)P 2 vesicles through its binding of the PIKfyve complex. Cellular and Molecular Life Sciences Heather Currinn, Benjamin Guscott, Zita Balklava, Alice
BILD7: Problem Set. 2. What did Chargaff discover and why was this important?
 BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
Electromagnetic characterization of magnetic steel alloys with respect to the temperature
 Electromagnetic characterization of magnetic steel alloys with respect to the temperature B Paya, P Teixeira To cite this version: B Paya, P Teixeira. Electromagnetic characterization of magnetic steel
Electromagnetic characterization of magnetic steel alloys with respect to the temperature B Paya, P Teixeira To cite this version: B Paya, P Teixeira. Electromagnetic characterization of magnetic steel