Spatio-Temporal Expression Patterns of Eight Epidermis-Specific Genes in the Ascidian Embryo
|
|
|
- Eugene Powers
- 6 years ago
- Views:
Transcription
1 Spatio-Temporal Expression Patterns of Eight Epidermis-Specific Genes in the Ascidian Embryo Author(s): Kouichi Ishida, Tatsuya Ueki, and Noriyuki Satoh Source: Zoological Science, 13(5): Published By: Zoological Society of Japan URL: BioOne ( is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne s Terms of Use, available at Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research.

2 ZOOLOGICAL SCIENCE 13: (1996) 1996 Zoological Society of Japan Spatio-Temporal Expression Patterns of Eight Epidermis-Specific Genes in the Ascidian Embryo Kouichi Ishida, Tatsuya Ueki 1 and Noriyuki Satoh Department of Zoology, Graduate School of Science, Kyoto University, Sakyo-ku, Kyoto , Japan ABSTRACT During embryogenesis of the ascidian Halocynthia roretzi, exactly eight-hundred epidermal cells are formed in the larva, and the lineage of the cells has been almost completely described. In the present study, we examined the spatio-temporal expression patterns of eight epidermis-specific genes which we already isolated. In situ hybridization of whole-mount specimens unambiguously demonstrated that the expression patterns of the eight genes were not identical, and that they were categorizable into several types. Transcripts of seven genes were restricted to presumptive epidermal cells, although transcripts of one gene were evident in the presumptive neural cells in addition to the presumptive epidermal cells. Therefore, most of the epidermis-specific genes in ascidian embryos are expressed in lineage-associated manner. We discuss these results in relation to the question of whether (a) epidermis-specific genes are expressed exclusively in presumptive epidermal cells prior to neural induction, or (b) the genes are first expressed in both epidermal precursors and precursors of the central nervous system, then the gene expression is downregulated in the latter after the completion of neural induction. Interestingly, cells of the anterior-most region as well as the dorsal midline of the tailbud embryo did not express most of the epidermis-specific genes, suggesting regional differences in the embryonic epidermis. Some other genes might be expressed in a complementary pattern in these regions. INTRODUCTION The epidermis of multicellular animals serves several important functions. Because the identity of individuals is achieved by this tissue, sensory machinery as well as defense machinery have evolved in association with epidermis. During embryogenesis of the ascidian Halocynthia roretzi, exactly 800 epidermal cells differentiate to cover the outermost part of the embryo (Nishida, 1987; reviewed by Satoh, 1994). The lineage of the epidermal cells has been defined almost completely (Conklin, 1905; Ortolani, 1962; Nishida, 1987; Nicol and Meinertzhagen, 1988). All the epidermal cells originate from the four cells of the animal half of the eight-cell embryo (the anterior a4.2 and the posterior b4.2 pairs of cells). The vegetal four blastomeres are not involved in the formation of epidermis. As early as the 16-cell stage, the developmental fate of a pair of animal blastomeres becomes restricted to epidermis. Five pairs of blastomeres of the 32-cell embryo and eleven pairs of the 64-cell embryo become the primordial epidermal cells (Nishida, 1987). Reflecting the early establishment of their developmental fate, presumptive epidermal cells show high potential for autonomous differentiation when they are isolated from early embryos (reviewed by Satoh, 1994). This autonomy 1 Present address: Department of Morphogenesis, Institute of Molecular Embryology and Genetics (IMEG), Kumamoto University School of Medicine, Honjo, Kumamoto 860, Japan. is dependent on prelocalized maternal factors (Nishida, 1994). We have already isolated and characterized several cdna clones that correspond to epidermis-specific genes of the ascidian embryo (Ueki et al., 1991, 1994; Ueki and Satoh, 1994, 1995). When cleavage of fertilized eggs of H. roretzi was blocked and the blocked eggs were cultured as one-celled embryos for about 30 hr, they developed features of differentiation of the epidermal cells only (e.g., Nishikata et al., 1988). Utilizing these characteristics, we isolated cdna clones for eight different epidermis-specific genes (Ueki et al., 1991; Ueki and Satoh, 1995). In the present study, using the cdna as probe, we examined the following questions as to the gene expression leading to epidermis differentiation in ascidian embryos: How is the expression of these epidermisspecific genes regulated spatially and temporally during the differentiation of epidermis? Are these epidermis-specific gene expressions regulated in the same manner, or differentially according to the gene or the region of the embryo? Are these genes really expressed exclusively in cells of epidermal lineage throughout the embryogenesis? Since the finding of the "organizer" by Spemann and Mangold (1924), the differentiation of epidermis in vertebrate embryos has been studied and discussed repeatedly in relation to the neural induction (e.g., Spemann, 1938; Slack, 1991). Transplantation of presumptive chordal mesoderm of an early amphibian embryo into a blastocoel of another embryo causes transdifferentiation of epidermal cells into neural cells. This induction, in turn, results
 are derived from ectoderm, and their early territories are continuous.")
3 700 K. ishida, T. Ueki and N. Satoh in the formation of secondary axis of the embryo. During normal embryogenesis, both the epidermis and the central nervous system (CNS) are derived from ectoderm, and their early territories are continuous. In this context, there is a question as to whether (a) epidermis-specific genes are expressed exclusively in differentiating epidermal cells before neural induction, or (b) the genes are first expressed in cells of presumptive epidermis and in those of CNS, and then the gene expression is downregulated in the latter after neural induction. MATERIALS AND METHODS Ascidian eggs and embryos Halocynthia roretzi was collected during the spawning season near the Asamushi Marine Biological Station of Tohoku University, Aomori, Japan. H. roretzi is a self-sterile hermaphrodite. Naturally spawned eggs were fertilized with a suspension of non-self sperm, and fertilized eggs were cultured in filtered seawater at about 13 C. Embryogenesis proceeded with a high degree of synchrony in various batches of eggs. At this temperature, the first cleavage took place about 2 hr after insemination, and embryos divided at approximately hourly intervals. They developed into gastrulae, neurulae, and earlytailbud stage embryos about 12, 15, and 20 hr after fertilization, respectively. Tadpole larvae hatched about 35 hr after fertilization. Eggs and embryos at appropriate stages were collected by lowspeed centrifugation and fixed for in situ hybridization. cdna probes forh.roretzi epidermis-specific genes Isolation and characterization of cdna clones for eight different epidermis-specific genes of H. roretzi embryos were reported previously (Ueki et al., 1991, 1994; Ueki and Satoh, 1994, 1995). The cdnas were inserted into the EcoRI site of the plasmid vector pbluescript II SK(+) or (-) (Stratagene, La Jolla, CA, USA). The length of the inserts ranged from 0.9 kb to 2.1 kb. Digoxigenin (DIG)-labeled sense and antisense probes were synthesized following the instructions from the suppliers of the kit (DIG RNA Labeling kit; Boehringer Mannheim, Heidelberg, Germany). Their final sizes were reduced to approximately 150 nucleotides by alkaline hydrolysis. In situ hybridization Whole-mount specimens were hybridized in situ using DIGlabeled antisense and sense RNA probes essentially according to the method described by Yasuo and Satoh (1994). Briefly, specimens were fixed in 4% paraformaldehyde in 0.1M MOPS buffer (ph 7.5), 0.5 M NaCl. After being thoroughly washed with PBT (phosphatebuffered saline containing 0.1% Tween 20), the specimens were treated with 2 µg/ml proteinase K (Sigma, St. Louis, MO, USA) in PBT for 30 min at 37 C, then post-fixed with 4% paraformaldehyde in PBT for 1 hr at room temperature. After a 1 -hr period of prehybridization at 42 C, the specimens were allowed to hybridize with the DIG-labeled probes at a concentration of 1 µg/ml for at least 16 hr at 42 C. After hybridization, the hybridization solution was gradually replaced by PBT, then the specimens were digested with 20 µg/ml RNase A (Sigma). Thereafter, the samples were incubated for 1 hr with 500 µlanti-digalkaline phosphatase conjugate, and color was developed according to the Boehringer protocol. After dehydration, some of the specimens were cleared by placing them in a 1:2 mixture (v/v) of benzyl alcohol and benzyl benzoate. RESULTS With the aid of in situ hybridization of whole-mount specimens, we carefully examined the distribution of transcripts of each of the eight epidermis-specific genes in H. roretzi embryos. In most cases of whole-mount in situ hybridization of ascidian embryos, signals are first detected in the nucleus of certain blastomeres, then the signals distribute over the entire cytoplasm (e.g., Yasuo and Satoh, 1993; Satou et al., 1995). This situation enables us to judge unambiguously which blastomeres of early embryos express the gene and which do not. Of the eight different epidermis-specific genes, HrEpiA, HrEpiB, HrEpiE, HrEpiF, HrEpiG and HrEpiH are exclusively zygotic, whereas HrEpiC and HrEpiD are both maternal and zygotic (Ueki et al., 1991; Ueki and Satoh, 1995). In the present study, we examined the zygotic expression of these genes. Controls hybridized with sense probes did not show signals above the background (data not shown). Lineage of cells of epidermis and CNS In order to help the reader's understanding of the spatial distribution of epidermis-specific gene transcripts, we briefly describe here the lineages of epidermis and CNS of the ascidian embryo. The lineages of epidermis and CNS are well described (Conklin, 1905; Ortolani, 1962; Nishida, 1987; Nicol and Meinertzhagen, 1988). The ascidian embryo is bilaterally symmetrical, the first cleavage plane being only a plane of symmetry. The gene expression is usually detected symmetrically in a pair of right and left blastomeres. For example, in the following description, we used the a8.25 cells instead of a pair of right a8.25 and left a8.25 cells. As shown in Fig. 1, the epidermis is derived entirely from blastomeres of the animal hemisphere of early embryos. As early as the 16-cell stage, the b5.4 cells have been restricted to give rise to epidermis (Fig. 1A). Through its derivatives of the b6.7 and b6.8 cells of the 32-cell embryo (Fig. 1B), then through the b7.13, b7.14, b7.15 and b7.16 cells of the 64-cell embryo (Fig. 1C), the b5.4 cells give rise to 256 epidermal cells of the ventral part of the tail of the mid-tailbud embryo, because each of the b7.13, b7.14, b7.15 and b7.16 cells give rise to a clone of 64 epidermal cells (Nishida, 1987; Fig. 1H). The a6.6, a6.8 and b6.6 cells become restricted to epidermis at the 32-cell stage. The first two pairs of cells give rise to a total of 256 epidermal cells of the lateral and ventral regions of the head of the mid-tailbud embryo after six subsequent divisions, whereas the b6.6 cells form 128 epidermal cells of the lateral part of the tail (a7.11, a7.12, a7.15, a7.16, b7.11, and b7.12 in Fig. 1H). At the 64-cell stage, the a7.14 cells also become restricted to give rise to 64 epidermal cells of the dorsal part of the head (Fig. 1H). Epidermis of the dorsal part of the trunk and tail is derived from the b8.18 and b8.20 cells (Fig. 1H), which are derivatives of the b5.3, b6.5, b7.9, and b7.10 cells. Each of these cells forms 32 epidermal cells. In addition, 32 epidermal cells of the anterior dorsal part of the head are derived from the a8.26 cells (Fig. 1H). Furthermore, the adhesive organ of the anterior-most region of the embryo is derived from the a8.18 and a8.20 cells (Fig. 1H). Cells constructing the CNS are derived from a7.9, a7.10, a7.13 (brain lineage), b7.9 (spinal cord lineage) and b7.10 (brain stem and spinal cord lineage) cells at the 64-cell stage (Fig. 1C), and a8.17, a8.19, a8.25 (brain lineage), b8.17 (spinal
.")

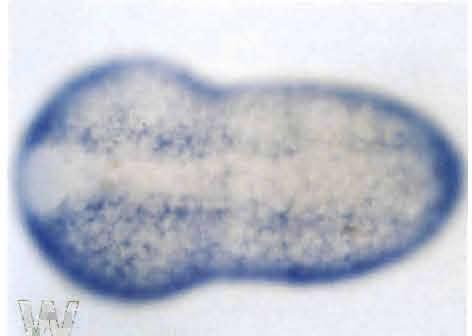
4 Epidermis-Specific Gene Expression 701 cord lineage) and b8.19 (brain stem and spinal cord lineage) at the 110-cell stage (Fig. 1D, E) in addition to A-line-derived neuronal cells. Spatial distribution of HrEpiA transcripts Northern blot analysis showed that the HrEpiA transcript is about 3.0 kb, and that HrEpiA expression is zygotic (Ueki et al., 1991; Ueki and Satoh, 1995). The transcript is first detected at the gastrula stage. The amount of the mrnas increases dramatically at the neurula and tailbud stages. However, the transcript is no longer detectable after the beginning of metamorphosis. Analyses of in situ hybridization demonstrated that the first distinct signals were detected at the early gastrula stage (Fig. 2A). At this stage, hybridization signals were evident in nuclei of the b8.25 and b8.27 primordial epidermal cells at the posterior-most region of the embryo (also see Fig. 1D, E). During gastrulation, signals became evident in many cells of the posterior-animal region of the embryo, and at the midgastrula stage, most of the b-line primordial epidermal cells showed hybridization signals (Fig. 2B). However, signals were not detected in the b8.17, b8.18, b8.19, b8.20, b-9.47, and b9.48 cells at both sides of the embryo, although the b8.18, b8.20, b9.47 and b9.48 cells are of epidermal lineage (the b8.17 cells are of spinal cord and muscle lineages, and the b8.19 cells are of neuronal lineage). At the neural-plate stage, signals were evident in the posterior epidermal cells (Fig. 2C, D). Although hybridization signals were not detected in a-line cells until the neural-plate stage, a pair of clusters consisting of about four cells (presumably descendants of the a7.14 cells) began to show signals at the neurula stage (Fig. 2E). It was noticed that the expression of HrEpiA was not detected in presumably one row of epidermal cells located most inside around the neural folds (Fig. 2E; compare with Fig. 4F). These cells seem to form the dorsal midline of the tail at the earlytailbud stage. As development proceeded, signals became evident in many a-line epidermal cells (Fig. 2F, G). However, there were cells without signals in the dorsal midline of the a- derived trunk region and the oblique line alongside it (Fig. 2F). In addition, the transcript began to disappear from epidermal cells at the midline of the ventral side of the embryo (Fig. 2G). Embryos at the early-tailbud stage showed a characteristic distribution pattern of HrEpiA transcripts (Fig. 2H, I). Signals were evident in almost all of epidermal cells except for the following three regions; the anterior-most region and about a line of cells at the boundary between the cells that started to express HrEpiA earlier among the a-line cells at the neurula stage and those that began to express the gene later; secondly, the dorsal midline of the trunk and tail region, which are Fig. 1. Diagram illustrating the lineage (A-G) and clonal organization (H) of epidermal cells during embryogenesis of the ascidian Halocynthia roretzi. (A) A 16-cell stage embryo, animal pole view; (B) a 32-cell stage embryo, animal pole view; (C) a 76-cell stage embryo, animal pole view (the pattern of the animal blastomeres of the 76-cell stage embryo is the same as that of the 64-cell stage embryo); (D, E) a 110-cell stage embryo, viewed from (D) the animal pole and (E) right side; and (F, G) a mid-gastrula, viewed from (F) the animal pole and (G) right side. Anterior is up. Blastomeres of epidermal, neuronal (shaded) and other lineages are shown by their nomenclatures. (H) The clonal organization of the epidermal cells of a tailbud embryo, with domains of ceils derived from epidermis-restricted blastomeres at the 64-cell and 110-cell stages. The domains derived from the a8.18 and a8.20 cells develop into palps of the larva. See the text for details. Based on description by Nishida (1987).


 An early gastrula viewed from the")
.")

 at the posterior-most region")

 A mid-gastrula viewed from the")



.")




.")

 the dorsal")


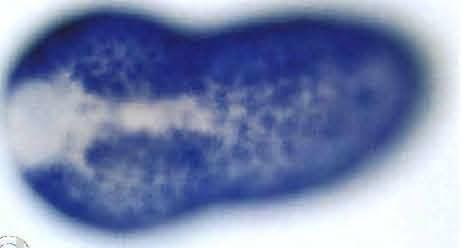
5 702 K. Ishida, T. Ueki and N. Satoh Fig. 2. Spatial distribution of HrEpiA transcripts, as revealed by whole-mount in situ hybridization with a digoxigenin-labeled antisense probe. (A) An early gastrula viewed from the vegetal pole (future dorsal side of the embryo). Hybridization signals are seen in nuclei of two pairs (b8.25 and b8.27) of the primordial epidermal cells (arrows) at the posterior-most region of the embryo. Bp, blastopore. Scale bar represents 50 µm for all panels. Anterior is up for all panels. (B) A mid-gastrula viewed from the animal pole. Signals are found in many primordial epidermal cells of the posterior region of the embryo. (C, D) Neural-plate stage embryos, animal side view. Focus is on the ventral surface (C) and the dorsal surface (D). Signals are seen in the posterior epidermal cells. (E) A neurula, dorsal view. A pair of clusters of a-line epidermal cells began to express the gene at the anterior side of the embryo (arrowheads). Signals cannot be seen in presumably one row of epidermal cells located most inside around the neural folds (arrows). The transcripts begin to disappear from epidermal cells at the posterior midline of the embryo. (F, G) An initial-tailbud stage embryo viewed from (F) the dorsal and (G) ventral side. A characteristic pattern of signal distribution is evident; signals are seen in the epidermal cells except those of the midline and the anterior-most regions of the embryo as well as the anterior ring (arrows in F). (H, I) An early-tailbud stage embryo viewed from (H) the dorsal and (I) ventral side. Distribution of signals is essentially identical to that of the initial tailbud embryo.
; thirdly, the ventral midline of the tail region, which are descendants of the b7.14 and b7.16 cells.")
6 Epidermis-Specific Gene Expression 703 descendants of the a7.14 and a8.26 (both are the trunk region, see also Fig. 1F), and the b8.18 and b8.20 ceils (both are the tail region); thirdly, the ventral midline of the tail region, which are descendants of the b7.14 and b7.16 cells. This distribution pattern of HrEpiA transcripts was retained at the middle-tailbud stage. However, signals of the cells in the ventral midline of the trunk region disappeared, and two patches of signals became distinct which were located horizontally below the ring of signals around the neuropore in the anterior-most region (data not shown). Therefore, HrEpiA is expressed in cells of differentiating epidermis but not in cells of the CNS. However, the gene was not expressed in epidermal cells of the midline of the tailbud embryo. Spatial distribution of HrEpiB transcripts HrEpiB shows a sequence similarity to UDP-glucose- 4-epimerases and 3β-hydroxysteroid dehydrogenase/ isomerases (Ueki et al., 1994). The transcript is about 1.2 kb. The activity of HrEpiB is also zygotic, and HrEpiB transcript is retained after metamorphosis by juveniles. HrEpiB transcript was first detected at the mid-gastrula stage (Fig. 3A). At this stage, hybridization signals were evident in many b-line primordial epidermal cells except for the b8.17, b8.18, b-8.19, b8.20, b9.47 and b9.48 cells, although signals were weak in the b9.59, b9.60, b9.63 and b9.64 cells (Fig. 3A; see also Fig. 1F, G). At the neurula stage, the expression was evident in b-line epidermal cells at the posterior part of the embryo. As in the case of HrEpiA, the expression of HrEpiB was undetectable in presumably one row of epidermal cells located most inside around the neural folds (Fig. 3B). After the neurula stage, in addition to b-line epidermal cells, a-line epidermal cells began to express the gene. In contrast to HrEpiA, however, all of the a-line lateral epidermal cells expressed HrEpiB (Fig. 3C). At the early-tailbud stage, the HrEpiB transcript was evident in the cells that occupied most of the outer surface, except for two regions; one was the anterior-most region of the embryo, descendants of the a8.18 and a8.20 cells, and the other was the anterior dorsal midline of the embryo, descendants of the a7.14, a8.26 and b8.20 cells (Fig. 3C). This distribution pattern of HrEpiB transcripts was retained at the middle-tailbud stage, except that signals of the cells in the ring around the neuropore disappeared discontinuously (data not shown). Therefore, HrEpiB was expressed in cells of differentiating epidermis but not in cells of the CNS, although the gene was not expressed later in cells of the anterior region and anterior midline of the tailbud embryo. Spatial distribution of HrEpiC transcripts The HrEpiC transcript is about 1.9 kb (Ueki et al., 1991; Ueki and Satoh, 1995). The gene expression is both maternal and zygotic, although HrEpiC mrna becomes undetectable soon after the beginning of metamorphosis. The pattern of appearance and distribution of HrEpiC transcripts differs from that of HrEpiA and HrEpiB. In situ hybridization demonstrated that hybridization signals were first detected at the 64-cell stage in all of the animal (a-line and b- line) blastomeres except for the a7.9, a7.10, b7.9 and b7.10 cells (Fig. 4A). The a7.9 cells give rise to brain and palps, a7.10 to brain, primordial pharynx and palps, b7.9 to epidermis, spinal cord, muscle and endodermal strand, and b7.10 to epidermis, brain stem, spinal cord and muscle, respectively (Nishida, 1987). The a7.13 cells expressed HrEpiC, although its developmental fate was yet not restricted to epidermis. At the 110-cell stage, the a7.13 cells divide into the a8.25 and a8.26 cells. The former gives rise to brain and pigment cells, and the latter is restricted to give rise to epidermis. The expression of HrEpiC was evident in the a8.26 cells but not in the a8.25 cells. At the 110-cell and mid-gastrula stages, most of the a- line cells showed signals, except for the a and a8.25 cells (the neural lineage and the palps lineage) and most of the b-line cells except for the b cells (Fig. 4B, C). During neurulation, HrEp/C-expressing epidermal cells of the ventral side extended to cover the dorsal side (Fig. 4D-F), but the signals were not detected in cells of the anterior-dorsal midline and anterior-most region of the embryo (Fig. 4G). At the earlytailbud stage, signals were evident in all of the epidermal cells except for those of the anterior-most region and anterior dorsal midline of the embryo (Fig. 4H, I). At the middle-tailbud stage, signals of cells in the dorsal midline became evident, although signals in the anterior-most region were still missing (data not shown). Therefore, HrEpiCis expressed in cells of differentiating epidermis but not in cells of the CNS. Although the a7.13 cells (containing both the epidermal and neuronal lineage) express the gene, the transcript was evident only in its epidermal descendants, not in its neural descendants. In addition, the gene expression was not detected in cells of the anterior-most region and anterior-dorsal midline of the tailbud embryo. Spatial distribution of HrEpiD transcripts HrEpiD is an ascidian homologue of mammalian and yeast SEC61 (Ueki and Satoh, 1994). Northern blot analysis shows that the HrEpiD transcript is about 2.3 kb. The HrEpiD expression is both maternal and zygotic, and a considerable amount of HrEpiD transcripts are retained by larvae and metamorphosed juveniles. As shown in Fig. 3D-G, the spatial expression pattern of HrEpiD resembles that of HrEpiB, although signals were very weak in gastrulae (Fig. 3D). HrEpiD transcript was first detected at the mid-gastrula stage in many b-line primordial epidermal cells with the possible exception of the b , b9.47 and b9.48 cells (Fig. 3D). At the neurula stage, in addition to b-line epidermal cells, a-line epidermal cells began to express the gene (Fig. 3F). In contrast to HrEpiC, however, cells of the anterior-most region also showed signals (Fig. 3F). At the earlytailbud stage, HrEpiD transcript was evident in cells that occupied most of the outer surface, except for the cranial, neuropore-like region of the embryo (Fig. 3G). This distribution pattern of HrEpiD transcripts was retained at the middle-tailbud



7 704 K. Ishida, T. Ueki and N. Satoh

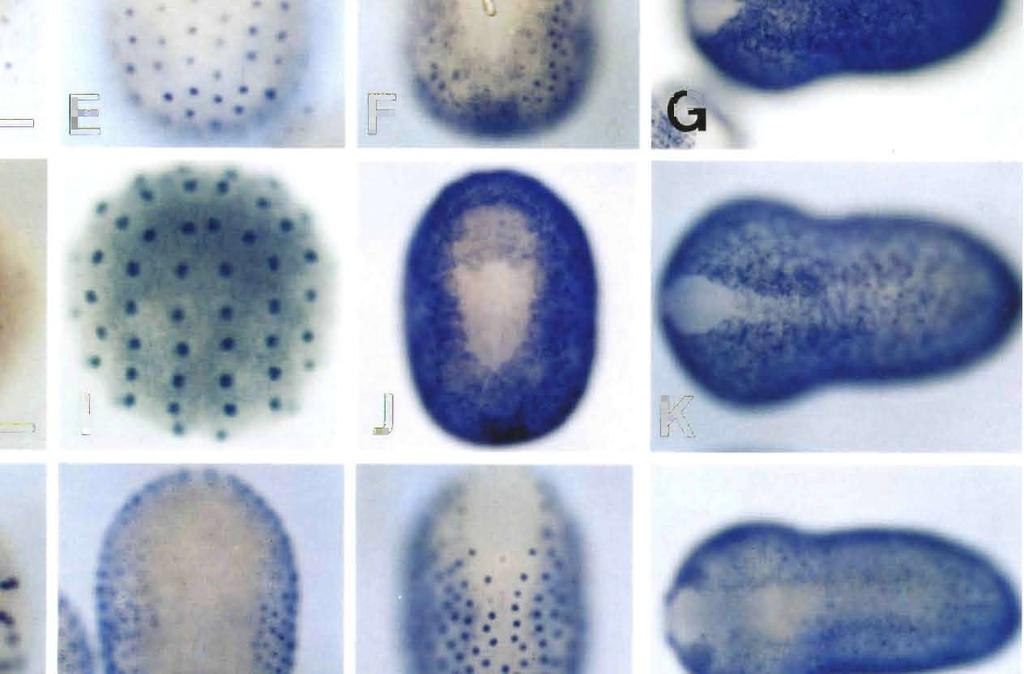
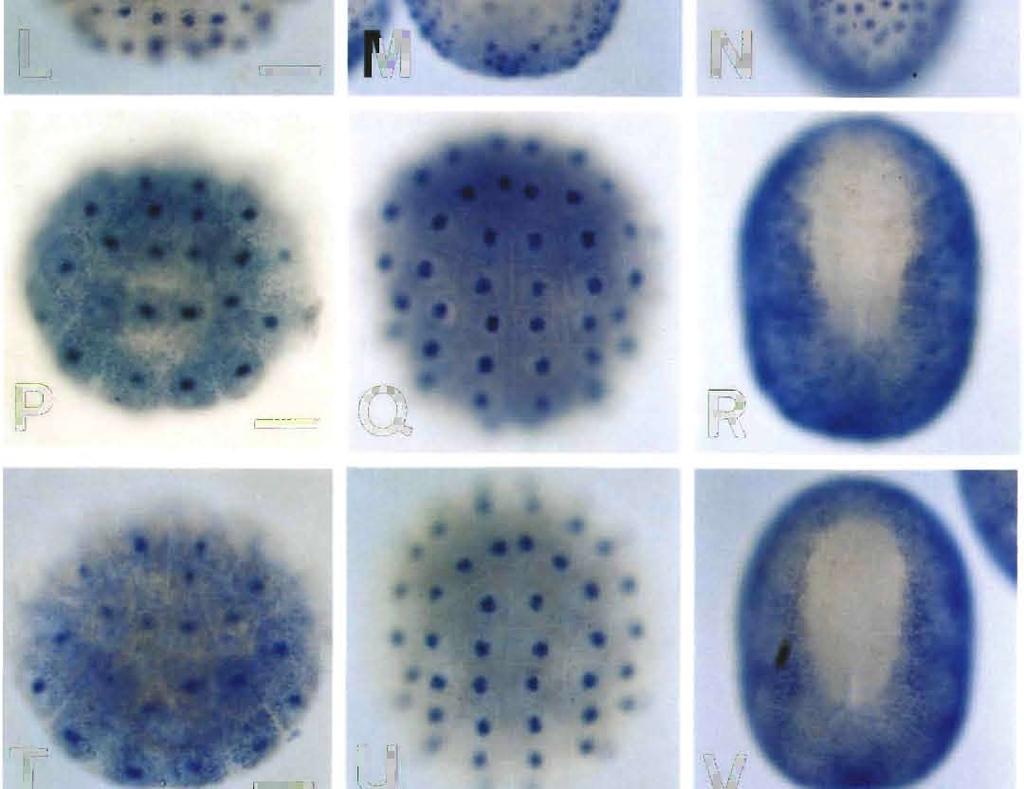
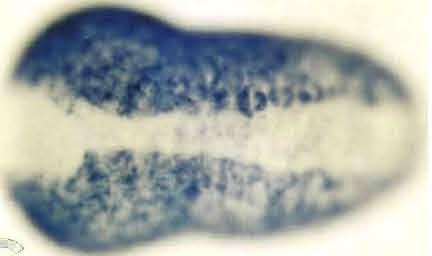
8 Epidermis-Specific Gene Expression 705 stage, i.e., signals were evident in almost all the epidermal cells except for the small neuropore region (data not shown). Therefore, HrEpiD was also not expressed in cells of the CNS. Spatial distribution of HrEpiE transcripts HrEpiE exhibits two different sizes of transcripts, about 2.3 and 3.0 kb in length, respectively (Ueki and Satoh, 1995). The HrEpiE expression is zygotic, and may no longer be detectable after the beginning of metamorphosis. An early phase of the appearance of HrEpiE transcripts resembles that of HrEpiC (Fig. 3H, I). Hybridization signals were first detected at the 76-cell stage in all of the animal (aline and b-line) blastomeres (Fig. 3H), including the a7.9, a7.10 and a7.13 cells, these three containing the neural lineage. In contrast to HrEpiC, HrEpiE transcripts were evident in cells of the neural lineage and the palps lineage (the a and a8.25 cells) at the 110-cell stage. During gastrulation and neurulation, HrEp/E-expressing epidermal cells of the ventral side extended over the dorsal side (Fig. 3I, J). At the earlytailbud stage, signals were evident in all of the epidermal cells except for those of the anterior, neuropore-like region and dorsal midline of the embryo (Fig. 3K). The signals in cells of the brain lineage disappeared by this stage. At the middletailbud stage, signals in almost all the epidermal cells including the dorsal midline of the embryo were evident although the signals were still missing in the small neuropore region (data not shown). Therefore, HrEpiE is expressed in cells of differentiating epidermis as well as presumptive brain cells. Spatial distribution of HrEpiF transcripts Northern blot analysis shows that the HrEpiF transcripts are about 4.8 and 10.0 kb in length (Ueki and Satoh, 1995). The HrEpiF expression is zygotic. In situ hybridization signals were detected in all of the b- line blastomeres except the b cells of the mid-gastrula (Fig. 3L). At the neurula stage, signals were evident in a-line epidermal cells although the signals in cells at the anteriormost region disappeared later (Fig. 3M). As in the case of HrEpiA, the expression of HrEpiF was not detected in one row of epidermal cells located most inside around the neural folds (Fig. 3M). These cells seem to form the future dorsal midline. At the initial-tailbud stage, signals were evident in cells of the dorsal midline (Fig. 3N), although the signals soon disappeared (Fig. 30). At the early-tailbud stage, signals were detected in all epidermal cells except for cells of the anteriormost region, neuropore-like region and dorsal midline of the embryo (Fig. 30). The pattern of HrEpiF expression at this stage resembles that of HrEpiA at the anterior part and the dorsal side of the embryo. This distribution pattern of HrEpiF transcripts was retained, except that the signals of the cells in the posterior-dorsal midline became evident (data not shown). Therefore, HrEpiF is expressed exclusively in cells of differentiating epidermal cells. Spatial distributionofhrepig transcripts HrEpiG encodes two different sizes of transcripts, about 2.9 and 3.1 kb in length, respectively (Ueki and Satoh, 1995). The HrEpiG expression is zygotic, and a considerable amount of HrEpiG transcripts may be retained by larvae and metamorphosed juveniles. In a previous study (Ueki and Satoh, 1995), we detected HrEpiG transcripts by faint Northern blot at the 64-cell stage. In the present study, however, first hybridization signals of HrEpiG were detected at the 76-cell stage in a-line blastomeres, except for the a7.9, a7.10 and a7.13 cells, and in b-line blastomeres except for the b7.9 and b7.10 cells (Fig. 3P). At the 110-cell stage, signals were found in a-line blastomeres except for the a8.17, a8.19 and a8.25 cells and in b-line blastomeres except for the b8.17 and b8.19 cells (Fig. 3Q). At the neurula stage, signals were evident in all of the epidermal cells (Fig. 3R), although signals in cells at the anterior-most region later disappeared. At the early-tailbud stage, signals were found in epidermal cells except for cells of the anterior-most region and dorsal midline of the embryo (Fig. 3S). This distribution pattern of HrEpiG transcripts was retained also at the middle-tailbud stage (data not shown). Therefore, HrEpiG is expressed in cells of differentiating epidermis but not in cells of the CNS. Spatial distribution of HrEpiH transcripts The HrEpiH transcripts are about 1.1 and 1.5 kb in length (Ueki and Satoh, 1995). The gene expression is zygotic, and a considerable amount of HrEpiH transcripts may be retained by larvae and metamorphosed juveniles. The distribution pattern of HrEpiH transcripts resembled that of HrEpiG. In the previous study (Ueki and Satoh, 1995), we detected the transcripts at the 64-cell stage faintly by Northern blot analysis. In the present study, however, first hybridization signals were detected at the 76-cell stage in a- line blastomeres except for the a7.9, a7.10 and a7.13 cells Fig. 3. Spatial distribution of transcripts of six other epidermis-specific genes in ascidian embryos, as revealed by whole-mount in situ hybridization. Scale bar represents 50 µm for all panels. Anterior is up for all panels except C, G, K, O, S and W in which anterior is left. (A-C) HrEpiB. (A) A mid-gastrula, animal pole view. (B) A neurula, dorsal view. (C) An early-tailbud stage embryo, dorsal view. (D-G) HrEpiD (D) A midgastrula, animal pole view. (E) A neural-plate stage embryo, ventral view. (F) A neurula, dorsal view. (G) An early-tailbud stage embryo, dorsal view. (H-K) HrEpiE. (H) A 76-cell embryo, animal pole view. (I) A 110-cell embryo, animal pole view. (J) A neurula, dorsal view. (K) An early-tailbud stage embryo, dorsal view. (L-O) HrEpiF. (L) A mid-gastrula, animal pole view. (M) A neurula, dorsal view. (N) An initial-tailbud stage embryo, dorsal view. (O) An early-tailbud stage embryo, dorsal view. (P-S) HrEpiG. (P) A 76-cell embryo, animal pole view. (Q) A 110- cell embryo, animal pole view. (R) A neurula, dorsal view. (S) An early-tailbud stage embryo, dorsal view. (T-W) HrEpiH. (T) A 76-cell embryo, animal pole view. (U) A 110-cell embryo, animal pole view. (V) A neurula, dorsal view. (W) An early-tailbud stage embryo, dorsal view.






 animal hemisphere.")



 the ventral and (E)")

 A neurula, dorsal view.")


.")


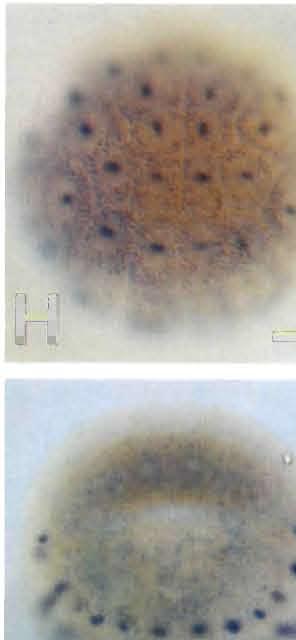
9 706 K. Ishida, T. Ueki and N. Satoh Fig. 4. Spatial distribution of HrEpiC transcripts, as revealed by whole-mount in situ hybridization with a digoxigenin-labeled antisense probe. (A) A 64-cell and (B) 110-cell stage embryo viewed from the animal pole (future ventral side). Hybridization signals are seen in nuclei of the presumptive as well as primordial epidermal cells at the anterior (a-line) and posterior (b-line) animal hemisphere. Scale bar represents 50 µm for all panels. Anterior is up for all panels. (C) An early gastrula viewed from the animal pole. Signals are evident in all of the primordial epidermal cells. (D, E) A neural-plate stage embryo, viewed from (D) the ventral and (E) dorsal side. Signals are seen in most of the epidermal cells. (F) A neurula, dorsal view. Epidermal cells exhibiting HrEpiC expression begin to cover the dorsal side of the embryo. HrEpiC expression is downregulated in cells of the anterior-most region at this stage (arrowhead). (G) An initial-tailbud stage embryo, dorsal viewed. Signals are seen in all the epidermal cells except those of the anterior-most region and anterior-dorsal midline of the embryo. (H, I) An early-tailbud embryo viewed from (H) the dorsal and (I) ventral side. Signals are seen in all of the epidermal cells except those of the anterior-most region and anterior-dorsal midline of the embryo.
. At the neurula stage, signals were evident in all of the epidermal cells (Fig.")
10 Epidermis-Specific Gene Expression 707 and in b-iine blastomeres except for the b7.9 and b7.10 cells (Fig. 3T). At the 110-cell stage, signals were found in a-line blastomeres except for the a8.17, a8.19 and a8.25 cells and in b-line blastomeres except for the b8.17 and b8.19 cells (Fig. 3U). At the neurula stage, signals were evident in all of the epidermal cells (Fig. 3V), and the signals in the cells of the anterior-most region were retained. At the early-tailbud stage, signals were found in epidermal cells except those of the cranial, neuropore-like region and the dorsal midline of the embryo (Fig. 3W). At the middle-tailbud stage, signals were evident in almost all of the epidermal cells except for the anterior-most region of the embryo (data not shown). Therefore, HrEpiH is also expressed in cells of differentiating epidermis but not in cells of the CNS. DISCUSSION Lineage-associated expression of epidermis-specific genes As shown in this study, most of these epidermis-specific genes are expressed exclusively in cells of epidermal lineage but not in cells of other lineages. Therefore, it is likely that the differentiation of epidermal cells in the ascidian embryo is achieved by the lineage-associated expression of the specific genes. However, one gene (HrEpiE) is expressed transiently in cells of the presumptive nervous system in addition to the cells of the presumptive epidermis. The eight epidermis-specific genes are categorized into two major groups based on the spatio-temporal expression patterns; the first group (HrEpiA, HrEpiB, HrEpiD, and HrEpiF) initiates their expression in b-line cells around the mid-gastrula stage and later in a-line cells around the neurula stage. In contrast, the second group (HrEpiC, HrEpiE, HrEpiG, and HrEpiH) starts their expression in almost all the a- and b-line cells of the animal hemisphere at the 76-cell stage. The expression patterns of the first group; differential regulation in epidermal lineage The first four genes, HrEpiA, HrEpiB, HrEpiD and HrEpiF, are expressed around the mid-gastrula stage only in b-line epidermal lineage cells. Their expression pattern seems to be regulated dependently upon their origins of blastomeres and simultaneously under a lineage-associated manner. These genes start their expression also in the a-line cells around the neurula stage. Thus, the temporal expression of these genes is differentially regulated between a- and b-line presumptive epidermal cells. It is an intriguing research subject to examine how the timing of the gene expression in certain blastomeres is determined dependently upon the origin of the blastomere. Although the significance of such differential expression is not known, a-line epidermal cells contribute exclusively to the epidermis of the trunk region, and b-line cells to that of the tail region. The expression pattern of the second group in relation to neural induction The four genes of the second group start their expression around the 76-cell stage in most of the cells in the animal hemisphere. At the 76-cell stage, the a7.9, a7.10, and a7.13 cells have both epidermal and neuronal fates (Fig. 1A), and at the 110-cell stage, the a8.17, a8.19, and a8.25 cells inherit the neuronal fate, whereas the a8.18, a8.20, and a8.26 cells inherit the epidermal and palps fate (Fig. 1B, C). As shown in this study, the distributions of HrEpiG and HrEpiH transcripts are restricted to cells of epidermal lineage at the 76-cell stage, and so is the distribution of HrEpiC transcripts, except for the distribution to the a7.13 cells. At the 110-cell stage, these three transcripts are evident exclusively in cells of epidermal lineage. Neural induction, at the least, has not been completed yet at the 110-cell stage (Nishida, 1991). Therefore, the expressions of these three genes are restricted entirely to epidermal lineage prior to the completion of neural induction. This implies that the presumptive epidermal cells and the presumptive neuronal cells have intrinsic differences already at the time before the completion of the neural induction. This is consistent with the observation that the brain-precursor cells isolated at the 110- cell stage failed to develop epidermis-specific features (Nishida, 1991). In contrast to the three genes mentioned above, HrEpiE is expressed in both cells of the epidermal and neuronal lineages from the 76-cell stage up to the neurula stage. This mode of gene expression is not interpreted as the lineageassociated mode of gene expression, as was observed in HrEpiC, HrEpiG and HrEpiH. The expression pattern of HrEpiE implies that the presumptive epidermal cells and the presumptive neuronal cells share some common characteristics around the 110-cell stage, and after neural induction the cells of neuronal lineage lose such characteristics. However, it is not known whether the loss of such characteristics is caused by the neural induction itself or by the intrinsic difference between cells of the neuronal and epidermal lineages that has already been established at the 110-cell stage, or by both. The isolated and cleavage-arrested a4.2 blastomere develops epidermal features, but in contact with A4.1 blastomere or treated with subtilisin, it develops neural features as a consequence of neural induction (reviewed in Okamura et al.,1993). Under such conditions, the inward-rectifier K + channel, which seems to be linked with epidermal differentiation in the above-mentioned system, is first expressed in a4.2 but thereafter suppressed by neural induction (Okamura and Takahashi, 1993). Therefore, the temporal expression pattern of this K + channel appears to be similar to that of HrEpiE, although the results of isolated and cleavage-arrested blastomeres do not always reflect the correct internal state of the cells in normal embryos. Spatial distribution pattern of epidermis-specific genes does not always coincide with temporal expression pattern On the basis of patterns of their temporal expression, the eight epidermis-specific genes of Halocynthia embryos are divided into four types (Table 1; Ueki and Satoh, 1995). Expression of the type I (HrEpiA, HrEpiE and HrEpiF) and type II (HrEpiB, HrEpiG and HrEpiH) is zygotic. In the case of
11 708 K. Ishida, T. Ueki and N. Satoh Table 1. Summary of the spatial expression patterns of eight epidermisspecific genes in the early-tailbud embryo Genes Type* regions + 1 HrEpiD IV Spatial distribution HrEpiB HrEpiC HrEpiE HrEpiH HrEpiG II III II II HrEpiF I HrEpiA I * The Roman numerals represent the type of temporal expression patterns (Ueki et al. 1995; see the text for details). + See Fig. 5 for six regions of the early-tailbud embryo. Fig. 5. Six regions in the epidermis of the early-tailbud embryo. The neuropore does not close completely at this stage, so the anteriorupper region (shaded area) is not covered with the epidermis yet. See Table 1. the types III (HrEpiC) and IV (HrEpiD), their expression is controlled both maternally and zygotically. In the case of the types I and III, transcripts become undetectable soon after the metamorphosis begins. Therefore, the activity of these genes is required for the formation of larval epidermis. In contrast to types I and III, mrnas of the types II and IV are retained throughout metamorphosis and by newly formed juveniles. Therefore, the activity of these genes is required not only for the formation of larval epidermis but also for the formation of juvenile epidermis. In another mode of classification, as mentioned before, the eight epidermis-specific genes are subdivided into two groups based on the spatio-temporal distribution patterns; four genes of the first group (HrEpiA, HrEpiB, HrEpiD, and HrEpiF) initiate their express in b-line cells around the mid-gastrula stage and later in a-line cells around the neurula stage. The other four genes of the second group (HrEpiC, HrEpiE, HrEpiG, and HrEpiH) start to express in almost all the a- and b-line cells of animal hemisphere. When we compared these spatiotemporal expression patterns with the temporal expression patterns (Table 1), we could not find any direct relationship between these two patterns. Furthermore, the spatial expression patterns of these eight genes at the early-tailbud stage are also divisible into six groups, as discussed below (Table 1). In this case, again, we could not find any relationship among these three modes of classification. Regional difference in epidermal cells One of the characteristic features of the eight different epidermis-specific genes in the ascidian embryo, shown in this study, is that many of the genes are not expressed in the anterior-most region as well as in the dorsal midline of the tailbud embryo. In previous studies (Ueki et al., 1991, 1994), we did not detect an exact spatial expression pattern of the epidermis-specific genes, as revealed in the present study; that is, in the previous studies, hybridization signals were detected all over the outer surface of the tailbud embryo. This difference is mainly caused by probes we used, because in the previous studies we used a cdna probe containing both strands (Ueki et al., 1991, 1994; Ueki and Satoh, 1994), although we used the antisense RNA probe to detect the transcripts of experimental embryos (Ueki et al., 1994). Thus, in the present study we could obtain precise expression patterns of the eight epidermis-specific genes and detect regional differences in epidermal cells based on these distribution patterns. Based on their spatial expression patterns at the earlytailbud stage, the eight genes can be divided into six groups

12 Epidermis-Specific Gene Expression 709 (Table 1). In other words, the epidermis of early-tailbud embryos is partitioned into six regions according to the set of gene expression (Table 1; Fig. 5). These six regions are thought to have intrinsic differences from one another. Then, what structural differences would be there in these six regions? In the ascidian tadpole, several structures are known to be derived from epidermis. The adhesive organ is formed from region 3 (Fig. 5). This organ consists of triangular papillae which are enveloped by a single layer of flattened epithelial cells, except at the anterior tips. Regions 2 and 5 may form the dorsal and ventral fins, respectively. Furthermore, it is reported in several ascidian species that the epidermal sensory neurons lie within these regions and that their axons run in grooves in the bases of epidermal cells along the dorsal and ventral midline of the tail (Torrence and Cloney, 1982). However, these epidermal sensory neurons are located pairwise at somewhat irregular intervals. Therefore, it seems unreasonable to relate this structure directly with regions 2 and 5. in region 1, the existence of the epidermal sensory neurons are reported in Ciona intestinalis (Takamura, 1995). However, they occupy only a small part of region 1, suggesting no direct relationship between this structure and the region. In region 4, no specific structure is found as yet, although it might be possible that some novel structure will be found in this region in future. As shown in our present study, most of the epidermisspecific genes are not expressed in the cells of the dorsal midline and the anterior-most region of the early-tailbud embryo. This implies that some other genes, which may confer some characteristics on the cells of this region, are expressed in a complementary pattern there. Recently, Miya et al. (1996) isolated an ascidian homologue (HrBMPa) of vertebrate BMPs5-8, and they showed that the gene is expressed in the adhesive organ, midline of the anterior dorsal neuroectoderm and midline of both ventral and dorsal ectoderm. Therefore, the spatial expression of HrBMPa seems to compensate for the expression of epidermis-specific genes. These expression patterns should be focused on in future studies. ACKNOWLEDGMENTS We wish to thank Dr. Takaharu Numakunai and all members of the Asamushi Marine Biological Station of Tohoku University for their hospitality. This research was supported in part by a Grant-in-Aid for Specially Promoted Research (No ) from the Ministry of Education, Science, Sports and Culture of Japan to N.S. REFERENCES Conklin EG (1905) The organization and cell lineage of the ascidian egg. J Acad Nat Sci Philadelphia 13: Miya T, Morita K, Ueno N, Satoh N (1996) An ascidian homologue of vertebrate BMPs5-8 is expressed in the midline of the anterior neuroectoderm and in the midline of the ventral epidermis of the embryo. Mech Dev 57: Nicol DR, Meinertzhagen IA (1988) Development of the central nervous system of the larva of the ascidian, Ciona intestinalisl.ii. Neural plate morphogenesis and cell lineage during neurulation. Dev Biol 130: Nishida H (1987) Cell lineage analysis in ascidian embryos by intracellular injection of a tracer enzyme. III. Up to the tissue restricted stage. Dev Biol 121: Nishida H (1991) Induction of brain and sensory pigment cells in the ascidian embryo analyzed by experiments with isolated blastomeres. Development 112: Nishida H (1994) Localization of egg cytoplasm that promotes differentiation to epidermis in embryos of the ascidian Halocynthia roretzi. Development 120: Nishikata T, Mita-Miyazawa I, Satoh N (1988) Differentiation expression in blastomeres of cleavage-arrested embryos of the ascidian Halocynthia roretzi. Dev Growth Differ 30: Okamura Y, Okado H, Takahashi K (1993) The ascidian embryo as a prototype of vertebrate neurogenesis. BioEssays 15: Okamura Y, Takahashi K (1993) Neural induction suppresses early expression of the inward-rectifier K + channel in the ascidian blastomere. J Physiol (Lond) 463: Ortolani G (1962) Territorio presuntivo del sistema nervaso nelle larve di ascidie. Acta Embryol Morphol Exp 5: Satoh N (1994) Developmental Biology of Ascidians. Cambridge Univ Press, New York Satou Y, Kusakabe T, Araki I, Satoh N (1995) Timing of initiation of muscle-specific gene expression in the ascidian embryo precedes that of developmental fate restriction in lineage cells. Dev Growth Differ 37: Slack JMW (1991) From Eggs to Embryos. Cambridge Univ Press, Cambridge Spemann H (1938) Embryonic Development and Induction. Yale Univ Press, New Heaven Spemann H, Mangold H (1924) Über Induktion von Embryonalanlagen durch Implantation artfremder Organisatoren. Roux's Arch Entwicklungsmech 100: Takamura K (1995) Studies on development of the nervous system in the ascidian Ciona intestinalis. PhD Thesis, Kyoto Univ Torrence SA, Cloney RA (1982) Nervous system of ascidian larvae: caudal primary sensory neurons. Zoomorphology 99: Ueki T, Makabe KW, Satoh N (1991) Isolation of cdna clones for epidermis-specific genes of the ascidian embryo. Dev Growth Differ 33: Ueki T, Yoshida S, Marikawa Y, Satoh N (1994) Autonomy of expression of epidermis-specific genes in the ascidian embryo. Dev Biol 164: Ueki T, Satoh N (1994) An ascidian homolog of SEC61 is expressed predominantly in epidermal cells of the embryo. Dev Biol 165: Ueki T, Satoh N (1995) Sequence motifs shared by the 5' flanking regions of two epidermis-specific genes in the ascidian embryo. Dev Growth Differ 37: Yasuo H, Satoh N (1993) Function of vertebrate Tgene. Nature 364: Yasuo H, Satoh N (1994) An ascidian homolog of the mouse Brachyury (7) gene is expressed exclusively in notochord cells at the fate restricted stage. Dev Growth Differ 36: 9-18 (Received April 25, 1996 / Accepted May 21, 1996)
posterior end mark, a novel maternal gene encoding a localized factor in the
 Development 122, 2005-2012 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV5066 2005 posterior end mark, a novel maternal gene encoding a localized factor in the ascidian embryo
Development 122, 2005-2012 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV5066 2005 posterior end mark, a novel maternal gene encoding a localized factor in the ascidian embryo
On the a clock' mechanism determining the time of tissue-specific enzyme development during ascidian embryogenesis
 /. Embryo!, exp. Morph. Vol. 54, pp. 131-139, 1979 Printed in Great Britain Company of Biologists Limited 1979 On the a clock' mechanism determining the time of tissue-specific enzyme development during
/. Embryo!, exp. Morph. Vol. 54, pp. 131-139, 1979 Printed in Great Britain Company of Biologists Limited 1979 On the a clock' mechanism determining the time of tissue-specific enzyme development during
Temporal expression patterns of 39 Brachyurydownstream genes associated with notochord formation in the Ciona intestinalis embryo
 Develop. Growth Differ. (1999) 41, 657 664 Temporal expression patterns of 39 Brachyurydownstream genes associated with notochord formation in the Ciona intestinalis embryo Kohji Hotta, 1 * Hiroki Takahashi,
Develop. Growth Differ. (1999) 41, 657 664 Temporal expression patterns of 39 Brachyurydownstream genes associated with notochord formation in the Ciona intestinalis embryo Kohji Hotta, 1 * Hiroki Takahashi,
Maternal Control of GermLayer Formation in Xenopus
 Maternal Control of GermLayer Formation in Xenopus The zygotic genome is activated at the mid-blastula transition mid-blastula fertilized egg Xenopus gastrulae early-gastrula 7 hrs 10 hrs control not VP
Maternal Control of GermLayer Formation in Xenopus The zygotic genome is activated at the mid-blastula transition mid-blastula fertilized egg Xenopus gastrulae early-gastrula 7 hrs 10 hrs control not VP
Unicellular: Cells change function in response to a temporal plan, such as the cell cycle.
 Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11589 Supplementary Figure 1 Ciona intestinalis and Petromyzon marinus neural crest expression domain comparison. Cartoon shows dorsal views of Ciona mid gastrula (left) and Petromyzon
doi:10.1038/nature11589 Supplementary Figure 1 Ciona intestinalis and Petromyzon marinus neural crest expression domain comparison. Cartoon shows dorsal views of Ciona mid gastrula (left) and Petromyzon
Regionality of egg cytoplasm that promotes muscle differentiation in embryo of the ascidian, Halocynthia roretzi
 Development 116, 521-529 (1992) Printed in Great Britain The Company of Biologists Limited 1992 521 Regionality of egg cytoplasm that promotes muscle differentiation in embryo of the ascidian, Halocynthia
Development 116, 521-529 (1992) Printed in Great Britain The Company of Biologists Limited 1992 521 Regionality of egg cytoplasm that promotes muscle differentiation in embryo of the ascidian, Halocynthia
Early Development in Invertebrates
 Developmental Biology Biology 4361 Early Development in Invertebrates October 25, 2006 Early Development Overview Cleavage rapid cell divisions divisions of fertilized egg into many cells Gastrulation
Developmental Biology Biology 4361 Early Development in Invertebrates October 25, 2006 Early Development Overview Cleavage rapid cell divisions divisions of fertilized egg into many cells Gastrulation
Early specification of ascidian larval motor neurons
 Developmental Biology 278 (2005) 310 322 www.elsevier.com/locate/ydbio Early specification of ascidian larval motor neurons Yu Katsuyama a,b, *, Toshiaki Okada a,c, Jun Matsumoto a,d, Yukio Ohtsuka a,
Developmental Biology 278 (2005) 310 322 www.elsevier.com/locate/ydbio Early specification of ascidian larval motor neurons Yu Katsuyama a,b, *, Toshiaki Okada a,c, Jun Matsumoto a,d, Yukio Ohtsuka a,
Questions in developmental biology. Differentiation Morphogenesis Growth/apoptosis Reproduction Evolution Environmental integration
 Questions in developmental biology Differentiation Morphogenesis Growth/apoptosis Reproduction Evolution Environmental integration Representative cell types of a vertebrate zygote => embryo => adult differentiation
Questions in developmental biology Differentiation Morphogenesis Growth/apoptosis Reproduction Evolution Environmental integration Representative cell types of a vertebrate zygote => embryo => adult differentiation
An essential role of a FoxD gene in notochord induction in Ciona embryos
 Development 129, 3441-3453 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV5028 3441 An essential role of a FoxD gene in notochord induction in Ciona embryos Kaoru S. Imai*, Nori
Development 129, 3441-3453 (2002) Printed in Great Britain The Company of Biologists Limited 2002 DEV5028 3441 An essential role of a FoxD gene in notochord induction in Ciona embryos Kaoru S. Imai*, Nori
BIOLOGY - CLUTCH CH.32 - OVERVIEW OF ANIMALS.
 !! www.clutchprep.com Animals are multicellular, heterotrophic eukaryotes that feed by ingesting their food Most animals are diploid, and produce gametes produced directly by meiosis Animals lack cell
!! www.clutchprep.com Animals are multicellular, heterotrophic eukaryotes that feed by ingesting their food Most animals are diploid, and produce gametes produced directly by meiosis Animals lack cell
Neural development its all connected
 Neural development its all connected How do you build a complex nervous system? How do you build a complex nervous system? 1. Learn how tissue is instructed to become nervous system. Neural induction 2.
Neural development its all connected How do you build a complex nervous system? How do you build a complex nervous system? 1. Learn how tissue is instructed to become nervous system. Neural induction 2.
Developmental Biology Lecture Outlines
 Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018
 Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Developmental Biology 3230 Midterm Exam 1 March 2006
 Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Life Sciences For NET & SLET Exams Of UGC-CSIR. Section B and C. Volume-08. Contents A. BASIC CONCEPT OF DEVELOPMENT 1
 Section B and C Volume-08 Contents 5. DEVELOPMENTAL BIOLOGY A. BASIC CONCEPT OF DEVELOPMENT 1 B. GAMETOGENESIS, FERTILIZATION AND EARLY DEVELOPMENT 23 C. MORPHOGENESIS AND ORGANOGENESIS IN ANIMALS 91 0
Section B and C Volume-08 Contents 5. DEVELOPMENTAL BIOLOGY A. BASIC CONCEPT OF DEVELOPMENT 1 B. GAMETOGENESIS, FERTILIZATION AND EARLY DEVELOPMENT 23 C. MORPHOGENESIS AND ORGANOGENESIS IN ANIMALS 91 0
Question Set # 4 Answer Key 7.22 Nov. 2002
 Question Set # 4 Answer Key 7.22 Nov. 2002 1) A variety of reagents and approaches are frequently used by developmental biologists to understand the tissue interactions and molecular signaling pathways
Question Set # 4 Answer Key 7.22 Nov. 2002 1) A variety of reagents and approaches are frequently used by developmental biologists to understand the tissue interactions and molecular signaling pathways
Exam 1 ID#: October 4, 2007
 Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
8/23/2014. Introduction to Animal Diversity
 Introduction to Animal Diversity Chapter 32 Objectives List the characteristics that combine to define animals Summarize key events of the Paleozoic, Mesozoic, and Cenozoic eras Distinguish between the
Introduction to Animal Diversity Chapter 32 Objectives List the characteristics that combine to define animals Summarize key events of the Paleozoic, Mesozoic, and Cenozoic eras Distinguish between the
Cell lineage and determination of cell fate in ascidian embryos
 nt..j.de\', BiuJ.33: 197-111(19S9) 197 Review Cell lineage and determination of cell fate in ascidian embryos JUDTH M. VENUT and WilLAM R. JEFFERY' Center for Developmental Biology, Department of Zoology,
nt..j.de\', BiuJ.33: 197-111(19S9) 197 Review Cell lineage and determination of cell fate in ascidian embryos JUDTH M. VENUT and WilLAM R. JEFFERY' Center for Developmental Biology, Department of Zoology,
Chapter 10 Development and Differentiation
 Part III Organization of Cell Populations Chapter Since ancient times, people have wondered how organisms are formed during the developmental process, and many researchers have worked tirelessly in search
Part III Organization of Cell Populations Chapter Since ancient times, people have wondered how organisms are formed during the developmental process, and many researchers have worked tirelessly in search
Chapter 32. Objectives. Table of Contents. Characteristics. Characteristics, continued. Section 1 The Nature of Animals
 Introduction to Animals Table of Contents Objectives Identify four important characteristics of animals. List two kinds of tissues found only in animals. Explain how the first animals may have evolved
Introduction to Animals Table of Contents Objectives Identify four important characteristics of animals. List two kinds of tissues found only in animals. Explain how the first animals may have evolved
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its
 The Wnt/ -catenin signalling pathway and its") Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Role of Organizer Chages in Late Frog Embryos
 Ectoderm Germ Layer Frog Fate Map Frog Fate Map Role of Organizer Chages in Late Frog Embryos Organizer forms three distinct regions Notochord formation in chick Beta-catenin localization How does beta-catenin
Ectoderm Germ Layer Frog Fate Map Frog Fate Map Role of Organizer Chages in Late Frog Embryos Organizer forms three distinct regions Notochord formation in chick Beta-catenin localization How does beta-catenin
9/4/2015 INDUCTION CHAPTER 1. Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology. Fig 1.
 INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
Research article. Summary. Introduction. Kaoru S. Imai*, Kyosuke Hino*, Kasumi Yagi, Nori Satoh and Yutaka Satou
 Research article 4047 Gene expression profiles of transcription factors and signaling molecules in the ascidian embryo: towards a comprehensive understanding of gene networks Kaoru S. Imai*, Kyosuke Hino*,
Research article 4047 Gene expression profiles of transcription factors and signaling molecules in the ascidian embryo: towards a comprehensive understanding of gene networks Kaoru S. Imai*, Kyosuke Hino*,
Biosc 41 9/10 Announcements
 Biosc 41 9/10 Announcements v Genetics review: group problem sets Groups of 3-4 Correct answer presented to class = 2 pts extra credit Incorrect attempt = 1 pt extra credit v Lecture: Animal Body Plans
Biosc 41 9/10 Announcements v Genetics review: group problem sets Groups of 3-4 Correct answer presented to class = 2 pts extra credit Incorrect attempt = 1 pt extra credit v Lecture: Animal Body Plans
MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning
 MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning Learning Goals for Lecture 29 4.1 Describe the contributions of early developmental events in the embryo to the formation
MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning Learning Goals for Lecture 29 4.1 Describe the contributions of early developmental events in the embryo to the formation
Animal Diversity. Features shared by all animals. Animals are multicellular, heterotrophic eukaryotes with tissues that develop from embryonic layers
 Animal Diversity Animals are multicellular, heterotrophic eukaryotes with tissues that develop from embryonic layers Nutritional mode Ingest food and use enzymes in the body to digest Cell structure and
Animal Diversity Animals are multicellular, heterotrophic eukaryotes with tissues that develop from embryonic layers Nutritional mode Ingest food and use enzymes in the body to digest Cell structure and
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Outline. v Definition and major characteristics of animals v Dividing animals into groups based on: v Animal Phylogeny
 BIOSC 041 Overview of Animal Diversity: Animal Body Plans Reference: Chapter 32 Outline v Definition and major characteristics of animals v Dividing animals into groups based on: Body symmetry Tissues
BIOSC 041 Overview of Animal Diversity: Animal Body Plans Reference: Chapter 32 Outline v Definition and major characteristics of animals v Dividing animals into groups based on: Body symmetry Tissues
Dorsoventral patterning of the vertebrate neural tube is conserved in a protochordate
 Development 124, 2335-2344 (1997) Printed in Great Britain The Company of Biologists Limited 1997 DEV9537 2335 Dorsoventral patterning of the vertebrate neural tube is conserved in a protochordate Joseph
Development 124, 2335-2344 (1997) Printed in Great Britain The Company of Biologists Limited 1997 DEV9537 2335 Dorsoventral patterning of the vertebrate neural tube is conserved in a protochordate Joseph
Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8
 Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8 1. Inductive signaling is a hallmark of vertebrate and mammalian development. In early neural development, there are multiple signaling pathways
Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8 1. Inductive signaling is a hallmark of vertebrate and mammalian development. In early neural development, there are multiple signaling pathways
Animal Origins and Evolution
 Animal Origins and Evolution Common Features of Animals multicellular heterotrophic motile Sexual reproduction, embryo Evolution of Animals All animals are multicellular and heterotrophic, which means
Animal Origins and Evolution Common Features of Animals multicellular heterotrophic motile Sexual reproduction, embryo Evolution of Animals All animals are multicellular and heterotrophic, which means
Regulation of the Number of Cell Division Rounds by Tissue-Specific Transcription Factors and Cdk Inhibitor during Ascidian Embryogenesis
 Regulation of the Number of Cell Division Rounds by Tissue-Specific Transcription Factors and Cdk Inhibitor during Ascidian Embryogenesis Mami Kuwajima, Gaku Kumano, Hiroki Nishida* Department of Biological
Regulation of the Number of Cell Division Rounds by Tissue-Specific Transcription Factors and Cdk Inhibitor during Ascidian Embryogenesis Mami Kuwajima, Gaku Kumano, Hiroki Nishida* Department of Biological
Article. Localized PEM mrna and Protein Are Involved in Cleavage-Plane Orientation and Unequal Cell Divisions in Ascidians.
 Current Biology 17, 1014 1025, June 19, 2007 ª2007 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2007.05.047 Localized PEM mrna and Protein Are Involved in Cleavage-Plane Orientation and Unequal Cell
Current Biology 17, 1014 1025, June 19, 2007 ª2007 Elsevier Ltd All rights reserved DOI 10.1016/j.cub.2007.05.047 Localized PEM mrna and Protein Are Involved in Cleavage-Plane Orientation and Unequal Cell
PRACTICE EXAM. 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos.
 PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
Nature Biotechnology: doi: /nbt Supplementary Figure 1. Overexpression of YFP::GPR-1 in the germline.
 Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Supplementary Figure 1 Overexpression of YFP::GPR-1 in the germline. The pie-1 promoter and 3 utr were used to express yfp::gpr-1 in the germline. Expression levels from the yfp::gpr-1(cai 1.0)-expressing
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Novel Endostyle-Specific Genes in the Ascidian Ciona intestinalis
 ZOOLOGICAL SCIENCE 20: 1025 1030 (2003) 2003 Zoological Society of Japan Novel Endostyle-Specific Genes in the Ascidian Ciona intestinalis Akane Sasaki 1 *, Yuki Miyamoto 2, Yutaka Satou 1, Nori Satoh
ZOOLOGICAL SCIENCE 20: 1025 1030 (2003) 2003 Zoological Society of Japan Novel Endostyle-Specific Genes in the Ascidian Ciona intestinalis Akane Sasaki 1 *, Yuki Miyamoto 2, Yutaka Satou 1, Nori Satoh
Developmental Zoology. Ectodermal derivatives (ZOO ) Developmental Stages. Developmental Stages
 Developmental Stages. Developmental Stages") Developmental Zoology (ZOO 228.1.0) Ectodermal derivatives 1 Developmental Stages Ø Early Development Fertilization Cleavage Gastrulation Neurulation Ø Later Development Organogenesis Larval molts Metamorphosis
Developmental Zoology (ZOO 228.1.0) Ectodermal derivatives 1 Developmental Stages Ø Early Development Fertilization Cleavage Gastrulation Neurulation Ø Later Development Organogenesis Larval molts Metamorphosis
v Scientists have identified 1.3 million living species of animals v The definition of an animal
 Biosc 41 9/10 Announcements BIOSC 041 v Genetics review: group problem sets Groups of 3-4 Correct answer presented to class = 2 pts extra credit Incorrect attempt = 1 pt extra credit v Lecture: Animal
Biosc 41 9/10 Announcements BIOSC 041 v Genetics review: group problem sets Groups of 3-4 Correct answer presented to class = 2 pts extra credit Incorrect attempt = 1 pt extra credit v Lecture: Animal
Mutations affecting tail and notochord development in the ascidian Ciona
 Development 126, 3293-3301 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV6405 3293 Mutations affecting tail and notochord development in the ascidian Ciona savignyi Yuki Nakatani,
Development 126, 3293-3301 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV6405 3293 Mutations affecting tail and notochord development in the ascidian Ciona savignyi Yuki Nakatani,
Animal Diversity. Animals are multicellular, heterotrophic eukaryotes with tissues that develop from embryonic layers 9/20/2017
 Animal Diversity Chapter 32 Which of these organisms are animals? Animals are multicellular, heterotrophic eukaryotes with tissues that develop from embryonic layers Animals share the same: Nutritional
Animal Diversity Chapter 32 Which of these organisms are animals? Animals are multicellular, heterotrophic eukaryotes with tissues that develop from embryonic layers Animals share the same: Nutritional
Developmental Biology Biology 4361
 Developmental Biology Biology 4361 The Anatomical Tradition 2009 A hen is only an egg s way of making a new egg. Samuel Butler, 1885 The Anatomical Tradition - Overview What is developmental biology? How
Developmental Biology Biology 4361 The Anatomical Tradition 2009 A hen is only an egg s way of making a new egg. Samuel Butler, 1885 The Anatomical Tradition - Overview What is developmental biology? How
Development. 27 April 2017
 Development 27 April 2017 Development Development in multicellular organisms allow for cells and organ structures with specialized functions Development occurs at many points in the life cycle of an animal
Development 27 April 2017 Development Development in multicellular organisms allow for cells and organ structures with specialized functions Development occurs at many points in the life cycle of an animal
The Radiata-Bilateria split. Second branching in the evolutionary tree
 The Radiata-Bilateria split Second branching in the evolutionary tree Two very important characteristics are used to distinguish between the second bifurcation of metazoans Body symmetry Germinal layers
The Radiata-Bilateria split Second branching in the evolutionary tree Two very important characteristics are used to distinguish between the second bifurcation of metazoans Body symmetry Germinal layers
Chapter 32, 10 th edition Q1.Which characteristic below is shared by plants, fungi, and animals? ( Concept 32.1)
") Chapter 32, 10 th edition Q1.Which characteristic below is shared by plants, fungi, and animals? ( Concept 32.1) A) They are multicellular eukaryotes. B) They are heterotrophs. C) Their cells are supported
Chapter 32, 10 th edition Q1.Which characteristic below is shared by plants, fungi, and animals? ( Concept 32.1) A) They are multicellular eukaryotes. B) They are heterotrophs. C) Their cells are supported
Is maternal mrna a determinant of tissue-specific proteins in ascidian embryos?
 J. Embryol. exp. Morph. 97 Supplement, 1-14 (1986) Printed in Great Britain The Company of Biologists Limited 1986 Is maternal mrna a determinant of tissue-specific proteins in ascidian embryos? W. R.
J. Embryol. exp. Morph. 97 Supplement, 1-14 (1986) Printed in Great Britain The Company of Biologists Limited 1986 Is maternal mrna a determinant of tissue-specific proteins in ascidian embryos? W. R.
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis
 18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
Kingdom Animalia. Zoology the study of animals
 Kingdom Animalia Zoology the study of animals Summary Animals are multicellular and eukaryotic. consume and digest organic materials thereby being heterotrophs. Most are motile at some time in their lives.
Kingdom Animalia Zoology the study of animals Summary Animals are multicellular and eukaryotic. consume and digest organic materials thereby being heterotrophs. Most are motile at some time in their lives.
Sonic hedgehog (Shh) signalling in the rabbit embryo
 signalling in the rabbit embryo") Sonic hedgehog (Shh) signalling in the rabbit embryo In the first part of this thesis work the physical properties of cilia-driven leftward flow were characterised in the rabbit embryo. Since its discovery
Sonic hedgehog (Shh) signalling in the rabbit embryo In the first part of this thesis work the physical properties of cilia-driven leftward flow were characterised in the rabbit embryo. Since its discovery
Introduction to Animal Kingdom. Invertebrates and Vertebrates
 Introduction to Animal Kingdom Invertebrates and Vertebrates Introduction To Animals Vertebrate animal with a backbone. Invertebrate animal without a backbone; includes more than 95% of all animal species
Introduction to Animal Kingdom Invertebrates and Vertebrates Introduction To Animals Vertebrate animal with a backbone. Invertebrate animal without a backbone; includes more than 95% of all animal species
What Is an Animal? Section 25.1 Typical Animal Characteristics. I. Characteristics of Animals. Biology II Mrs. Michaelsen
 What Is an Animal? Section 25.1 Typical Animal Characteristics Biology II Mrs. Michaelsen I. Characteristics of Animals A. All animals are eukaryotic, multicellular, have ways of moving to reproduce, obtain
What Is an Animal? Section 25.1 Typical Animal Characteristics Biology II Mrs. Michaelsen I. Characteristics of Animals A. All animals are eukaryotic, multicellular, have ways of moving to reproduce, obtain
Biology 218, practise Exam 2, 2011
 Figure 3 The long-range effect of Sqt does not depend on the induction of the endogenous cyc or sqt genes. a, Design and predictions for the experiments shown in b-e. b-e, Single-cell injection of 4 pg
Figure 3 The long-range effect of Sqt does not depend on the induction of the endogenous cyc or sqt genes. a, Design and predictions for the experiments shown in b-e. b-e, Single-cell injection of 4 pg
Ascidian Laboratory Woods Hole Embryology Course, 2010
 Ascidian Laboratory Woods Hole Embryology Course, 2010 Robert W. Zeller rzeller@sciences.sdsu.edu Ascidians have a long history as an experimental model system for development. Edwin Grant Conklin followed
Ascidian Laboratory Woods Hole Embryology Course, 2010 Robert W. Zeller rzeller@sciences.sdsu.edu Ascidians have a long history as an experimental model system for development. Edwin Grant Conklin followed
!!!!!!!! DB3230 Midterm 2 12/13/2013 Name:
 1. (10 pts) Draw or describe the fate map of a late blastula stage sea urchin embryo. Draw or describe the corresponding fate map of the pluteus stage larva. Describe the sequence of gastrulation events
1. (10 pts) Draw or describe the fate map of a late blastula stage sea urchin embryo. Draw or describe the corresponding fate map of the pluteus stage larva. Describe the sequence of gastrulation events
Nature Neuroscience: doi: /nn.2662
 Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Supplementary Figure 1 Atlastin phylogeny and homology. (a) Maximum likelihood phylogenetic tree based on 18 Atlastin-1 sequences using the program Quicktree. Numbers at internal nodes correspond to bootstrap
Temporal regulation of the muscle gene cascade by Macho1 and Tbx6 transcription factors in Ciona intestinalis
 Research Article 2453 Temporal regulation of the muscle gene cascade by Macho1 and Tbx6 transcription factors in Ciona intestinalis Jamie E. Kugler 1, Stefan Gazdoiu 1, Izumi Oda-Ishii 1, Yale J. Passamaneck
Research Article 2453 Temporal regulation of the muscle gene cascade by Macho1 and Tbx6 transcription factors in Ciona intestinalis Jamie E. Kugler 1, Stefan Gazdoiu 1, Izumi Oda-Ishii 1, Yale J. Passamaneck
Lesson Overview. Gene Regulation and Expression. Lesson Overview Gene Regulation and Expression
 13.4 Gene Regulation and Expression THINK ABOUT IT Think of a library filled with how-to books. Would you ever need to use all of those books at the same time? Of course not. Now picture a tiny bacterium
13.4 Gene Regulation and Expression THINK ABOUT IT Think of a library filled with how-to books. Would you ever need to use all of those books at the same time? Of course not. Now picture a tiny bacterium
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
1. What are the three general areas of the developing vertebrate limb? 2. What embryonic regions contribute to the developing limb bud?
 Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Introduction to Animals
 Introduction to Animals Characteristics of Animals multicellular Except for sponges, animal cells are arranged into tissues. Tissues are necessary to produce organs and organ systems. Tissues, organs,
Introduction to Animals Characteristics of Animals multicellular Except for sponges, animal cells are arranged into tissues. Tissues are necessary to produce organs and organ systems. Tissues, organs,
An Introduction to Animal Diversity
 Chapter 32 An Introduction to Animal Diversity PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Chapter 32 An Introduction to Animal Diversity PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Principles of Experimental Embryology
 Biology 4361 Developmental Biology Principles of Experimental Embryology June 16, 2008 Overview What forces affect embryonic development? The embryonic environment: external and internal How do forces
Biology 4361 Developmental Biology Principles of Experimental Embryology June 16, 2008 Overview What forces affect embryonic development? The embryonic environment: external and internal How do forces
Expression of Hr-Erf Gene during Ascidian Embryogenesis
 Dev. Reprod. Vol. 17, No. 4, 389~397, December, 2013 ttp://dx.doi.org/10.12717/dr.2013.17.4.389 ISSN 1226-6752 (Print) ISSN 2287-7967 (Online) Expression of Hr-Erf Gene during Ascidian Embryogenesis *Jung
Dev. Reprod. Vol. 17, No. 4, 389~397, December, 2013 ttp://dx.doi.org/10.12717/dr.2013.17.4.389 ISSN 1226-6752 (Print) ISSN 2287-7967 (Online) Expression of Hr-Erf Gene during Ascidian Embryogenesis *Jung
THE NEUROBIOLOGY OF THE ASCIDIAN TADPOLE LARVA: Recent Developments in an Ancient Chordate
 Annu. Rev. Neurosci. 2004. 27:453 85 doi: 10.1146/annurev.neuro.27.070203.144255 Copyright c 2004 by Annual Reviews. All rights reserved THE NEUROBIOLOGY OF THE ASCIDIAN TADPOLE LARVA: Recent Developments
Annu. Rev. Neurosci. 2004. 27:453 85 doi: 10.1146/annurev.neuro.27.070203.144255 Copyright c 2004 by Annual Reviews. All rights reserved THE NEUROBIOLOGY OF THE ASCIDIAN TADPOLE LARVA: Recent Developments
Developmental processes Differential gene expression Introduction to determination The model organisms used to study developmental processes
 Date Title Topic(s) Learning Outcomes: Sept 28 Oct 3 1. What is developmental biology and why should we care? 2. What is so special about stem cells and gametes? Developmental processes Differential gene
Date Title Topic(s) Learning Outcomes: Sept 28 Oct 3 1. What is developmental biology and why should we care? 2. What is so special about stem cells and gametes? Developmental processes Differential gene
2. Fertilization activates the egg and bring together the nuclei of sperm and egg
 2. Fertilization activates the egg and bring together the nuclei of sperm and egg Sea urchins (what phylum?) are models for the study of the early development of deuterostomes (like us, right?). Sea urchin
2. Fertilization activates the egg and bring together the nuclei of sperm and egg Sea urchins (what phylum?) are models for the study of the early development of deuterostomes (like us, right?). Sea urchin
posterior end mark 2 (pem-2), pem-4, pem-5, and pem-6: Maternal Genes with Localized mrna in the Ascidian Embryo
, pem-4, pem-5, and pem-6: Maternal Genes with Localized mrna in the Ascidian Embryo") DEVELOPMENTAL BIOLOGY 192, 467 481 (1997) ARTICLE NO. DB978730 posterior end mark 2 (pem-2), pem-4, pem-5, and pem-6: Maternal Genes with Localized mrna in the Ascidian Embryo Yutaka Satou and Noriyuki
DEVELOPMENTAL BIOLOGY 192, 467 481 (1997) ARTICLE NO. DB978730 posterior end mark 2 (pem-2), pem-4, pem-5, and pem-6: Maternal Genes with Localized mrna in the Ascidian Embryo Yutaka Satou and Noriyuki
Brachyury downstream notochord differentiation in the ascidian embryo
 RESEARCH COMMUNICATION Brachyury downstream notochord differentiation in the ascidian embryo Hiroki Takahashi, 1,4 Kohji Hotta, 1,4 Albert Erives, 2,4 Anna Di Gregorio, 2 Robert W. Zeller, 3 Michael Levine,
RESEARCH COMMUNICATION Brachyury downstream notochord differentiation in the ascidian embryo Hiroki Takahashi, 1,4 Kohji Hotta, 1,4 Albert Erives, 2,4 Anna Di Gregorio, 2 Robert W. Zeller, 3 Michael Levine,
Designation of the Anterior/Posterior Axis in Pregastrula Xenopus laevis
 Developmental Biology 225, 37 58 (2000) doi:10.1006/dbio.2000.9803, available online at http://www.idealibrary.com on Designation of the Anterior/Posterior Axis in Pregastrula Xenopus laevis Mary Constance
Developmental Biology 225, 37 58 (2000) doi:10.1006/dbio.2000.9803, available online at http://www.idealibrary.com on Designation of the Anterior/Posterior Axis in Pregastrula Xenopus laevis Mary Constance
Chapter 32 Introduction to Animal Diversity. Copyright 2008 Pearson Education, Inc., publishing as Pearson Benjamin Cummings
 Chapter 32 Introduction to Animal Diversity Welcome to Your Kingdom The animal kingdom extends far beyond humans and other animals we may encounter 1.3 million living species of animals have been identified
Chapter 32 Introduction to Animal Diversity Welcome to Your Kingdom The animal kingdom extends far beyond humans and other animals we may encounter 1.3 million living species of animals have been identified
Direct activation by Ets and Zic is required for initial expression of the Brachyury gene in the ascidian notochord
 Developmental Biology 306 (2007) 870 882 www.elsevier.com/locate/ydbio Genomes & Developmental Control Direct activation by Ets and Zic is required for initial expression of the Brachyury gene in the ascidian
Developmental Biology 306 (2007) 870 882 www.elsevier.com/locate/ydbio Genomes & Developmental Control Direct activation by Ets and Zic is required for initial expression of the Brachyury gene in the ascidian
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Principles of Experimental Embryology
 Biology 4361 Developmental Biology Principles of Experimental Embryology September 19, 2006 Major Research Questions How do forces outside the embryo affect its development? (Environmental Developmental
Biology 4361 Developmental Biology Principles of Experimental Embryology September 19, 2006 Major Research Questions How do forces outside the embryo affect its development? (Environmental Developmental
Exam 2 ID#: November 9, 2006
 Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Drosophila melanogaster- Morphogen Gradient
 NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
Fish swimbladder: an excellent meso dermal inductor in primary embryonic induction
 /. Embryo/, exp. Morph. Vol 36,, pp. 315-30, 1976 315 Printed in Great Britain Fish swimbladder: an excellent meso dermal inductor in primary embryonic induction IZUMI KAWAKAMI 1 From the Department of
/. Embryo/, exp. Morph. Vol 36,, pp. 315-30, 1976 315 Printed in Great Britain Fish swimbladder: an excellent meso dermal inductor in primary embryonic induction IZUMI KAWAKAMI 1 From the Department of
Chapter 32 Introduction to Animal Diversity
 Chapter 32 Introduction to Animal Diversity Review: Biology 101 There are 3 domains: They are Archaea Bacteria Protista! Eukarya Endosymbiosis (proposed by Lynn Margulis) is a relationship between two
Chapter 32 Introduction to Animal Diversity Review: Biology 101 There are 3 domains: They are Archaea Bacteria Protista! Eukarya Endosymbiosis (proposed by Lynn Margulis) is a relationship between two
Caenorhabditis elegans
 Caenorhabditis elegans Why C. elegans? Sea urchins have told us much about embryogenesis. They are suited well for study in the lab; however, they do not tell us much about the genetics involved in embryogenesis.
Caenorhabditis elegans Why C. elegans? Sea urchins have told us much about embryogenesis. They are suited well for study in the lab; however, they do not tell us much about the genetics involved in embryogenesis.
Paraxial and Intermediate Mesoderm
 Biology 4361 Paraxial and Intermediate Mesoderm December 6, 2007 Mesoderm Formation Chick Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of
Biology 4361 Paraxial and Intermediate Mesoderm December 6, 2007 Mesoderm Formation Chick Major Mesoderm Lineages Mesodermal subdivisions are specified along a mediolateral axis by increasing amounts of
An Introduction to Animal Diversity
 Chapter 32 An Introduction to Animal Diversity PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Chapter 32 An Introduction to Animal Diversity PowerPoint Lecture Presentations for Biology Eighth Edition Neil Campbell and Jane Reece Lectures by Chris Romero, updated by Erin Barley with contributions
Comparison of the expression patterns of several sox genes between Oryzias latipes and Danio rerio
 Urun 1 Comparison of the expression patterns of several sox genes between Oryzias latipes and Danio rerio Fatma Rabia URUN ilkent University, nkara - TURKEY High mobility group domain containing transcription
Urun 1 Comparison of the expression patterns of several sox genes between Oryzias latipes and Danio rerio Fatma Rabia URUN ilkent University, nkara - TURKEY High mobility group domain containing transcription
An Introduction to Animal Diversity
 Chapter 32 An Introduction to Animal Diversity PowerPoint Lectures for Biology, Seventh Edition Neil Campbell and Jane Reece Lectures by Chris Romero Overview: Welcome to Your Kingdom The animal kingdom
Chapter 32 An Introduction to Animal Diversity PowerPoint Lectures for Biology, Seventh Edition Neil Campbell and Jane Reece Lectures by Chris Romero Overview: Welcome to Your Kingdom The animal kingdom
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Chapter 8-9 Intro to Animals. Image from:
 Chapter 8-9 Intro to Animals Image from: http://animaldiversity.ummz.umich.edu/index.html Zoology Definition: the scientific study of the behavior, structure, physiology, classification, and distribution
Chapter 8-9 Intro to Animals Image from: http://animaldiversity.ummz.umich.edu/index.html Zoology Definition: the scientific study of the behavior, structure, physiology, classification, and distribution
DEVELOPMENT. Clare Hudson*, Sonia Lotito and Hitoyoshi Yasuo
 Access the Development most First recent posted version epress online at on online http://dev.biologists.org/lookup/doi/10.1242/dev.002352 29 August publication 2007 as 10.1242/dev.002352 date 29 August
Access the Development most First recent posted version epress online at on online http://dev.biologists.org/lookup/doi/10.1242/dev.002352 29 August publication 2007 as 10.1242/dev.002352 date 29 August
Biology 376 Animal Development
 The stories are in every newspaper: cloning, stem cells, genetic engineering, in vitro fertilization, cancer therapies, organ regeneration, and protocols for prolonging our lifespan. In the past five years,
The stories are in every newspaper: cloning, stem cells, genetic engineering, in vitro fertilization, cancer therapies, organ regeneration, and protocols for prolonging our lifespan. In the past five years,
Recent studies have identified several signaling pathways and
 Evolutionary origins of the vertebrate heart: Specification of the cardiac lineage in Ciona intestinalis Brad Davidson* and Michael Levine Department of Molecular and Cell Biology, Division of Genetics
Evolutionary origins of the vertebrate heart: Specification of the cardiac lineage in Ciona intestinalis Brad Davidson* and Michael Levine Department of Molecular and Cell Biology, Division of Genetics
10/2/2015. Chapter 4. Determination and Differentiation. Neuroanatomical Diversity
 Chapter 4 Determination and Differentiation Neuroanatomical Diversity 1 Neurochemical diversity: another important aspect of neuronal fate Neurotransmitters and their receptors Excitatory Glutamate Acetylcholine
Chapter 4 Determination and Differentiation Neuroanatomical Diversity 1 Neurochemical diversity: another important aspect of neuronal fate Neurotransmitters and their receptors Excitatory Glutamate Acetylcholine
Drosophila Life Cycle
 Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:1.138/nature1237 a b retinol retinal RA OH RDH (retinol dehydrogenase) O H Raldh2 O R/R.6.4.2 (retinaldehyde dehydrogenase 2) RA retinal retinol..1.1 1 Concentration (nm)
SUPPLEMENTARY INFORMATION doi:1.138/nature1237 a b retinol retinal RA OH RDH (retinol dehydrogenase) O H Raldh2 O R/R.6.4.2 (retinaldehyde dehydrogenase 2) RA retinal retinol..1.1 1 Concentration (nm)
Subfamily-Specific Posttranscriptional Mechanism Underlies K Channel Expression in a Developing Neuronal Blastomere
 The Journal of Neuroscience, August 15, 1999, 19(16):6874 6886 Subfamily-Specific Posttranscriptional Mechanism Underlies K Channel Expression in a Developing Neuronal Blastomere Fumihito Ono, 1,2 You
The Journal of Neuroscience, August 15, 1999, 19(16):6874 6886 Subfamily-Specific Posttranscriptional Mechanism Underlies K Channel Expression in a Developing Neuronal Blastomere Fumihito Ono, 1,2 You
Divergent mechanisms specify chordate motoneurons: evidence from ascidians
 RESEARCH ARTICLE 1643 Development 138, 1643-1652 (2011) doi:10.1242/dev.055426 2011. Published by The Company of Biologists Ltd Divergent mechanisms specify chordate motoneurons: evidence from ascidians
RESEARCH ARTICLE 1643 Development 138, 1643-1652 (2011) doi:10.1242/dev.055426 2011. Published by The Company of Biologists Ltd Divergent mechanisms specify chordate motoneurons: evidence from ascidians
Developmental Biology
 Developmental Biology 344 (2010) 284 292 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology Macho-1 regulates unequal cell divisions
Developmental Biology 344 (2010) 284 292 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology Macho-1 regulates unequal cell divisions
Features of the Animal
 Features of the Animal Kingdom Bởi: OpenStaxCollege Even though members of the animal kingdom are incredibly diverse, animals share common features that distinguish them from organisms in other kingdoms.
Features of the Animal Kingdom Bởi: OpenStaxCollege Even though members of the animal kingdom are incredibly diverse, animals share common features that distinguish them from organisms in other kingdoms.