Dorsal Determinants in the Xenopus Egg Are Firmly Associated with the Vegetal Cortex and Behave like Activators of the Wnt Pathway
|
|
|
- Martin Wade
- 6 years ago
- Views:
Transcription
1 DEVELOPMENTAL BIOLOGY 191, (1997) ARTICLE NO. DB Dorsal Determinants in the Xenopus Egg Are Firmly Associated with the Vegetal Cortex and Behave like Activators of the Wnt Pathway Yusuke Marikawa, Yu Li, and Richard P. Elinson Department of Zoology, University of Toronto, 25 Harbord Street, Toronto M5S 3G5, Canada The Xenopus egg contains maternal dorsal determinants that are specifically located at the vegetal cortex. To study physical and functional properties of the dorsal determinants, we took advantage of the animal vegetal reversed embryo. The animal vegetal reversed embryo is produced by inversion of the fertilized egg, which results in formation of ectoderm and endoderm from the unpigmented and the pigmented halves, respectively [Neff et al. (1983). Dev. Biol. 97, ; Black and Gerhart (1985). Dev. Biol. 108, ]. We demonstrated by cytoplasmic transplantation that the dorsal activity was specifically localized to the unpigmented cortical cytoplasm of the inverted egg, which is segregated into the future ectodermal lineage. This result suggests that the dorsal determinants are associated with the unpigmented cortex and are not dislodged by the inversion. In addition, we found that two vegetally localized transcripts, Xcat2 and Vg1 mrnas, were present in the reversed animal pole of the inverted egg, suggesting their association with the unpigmented cortex. In order to compare the dorsal determinant activity with known dorsalizing molecules, we examined the expression pattern of Xnr3 and Siamois in the reversed embryo because these two genes are activated by the Wnt-pathway activators (Xwnt- 8, b-catenin, etc.) but not by other dorsalizing molecules (noggin, BVg1, etc.). Animal cap of the reversed embryo, which received the unpigmented cortex of the egg, expressed Xnr3 and Siamois. However, Mix.1, a marker expressed in endoderm and mesoderm in the normal embryo in response to mesodermal inducers, was not detected in the animal cap of the reversed embryo. In addition, we found that b-catenin protein accumulated in nuclei of unpigmented animal pole cells of the reversed embryo. These results suggest that the maternal dorsal determinants behave more similarly to the Wntpathway activators than noggin or BVg Academic Press INTRODUCTION Maternal information localized in particular regions of the egg cytoplasm is responsible for future determination of developmental fates in early embryos. In the Xenopus egg, the cytoplasmic factor that is responsible for determining dorsal structures is specifically located in the vegetal cortex. Microinjection of vegetal cortical cytoplasm into ventral blastomeres of a 16-cell recipient embryo generates a complete secondary dorsal axis (Fujisue et al., 1993; Holo- wacz and Elinson, 1993). Removal of the cortical cytoplasm from the vegetal pole of the early 1-cell embryo results in a dorsal deficiency (Kikkawa et al., 1996; Sakai, 1996). The dorsal determinants in the vegetal cortex alter the response of animal cap cells to mesoderm inducers and are unlikely to be mesoderm inducers (Holowacz and Elinson, 1995), although the molecular nature of the dorsal determinants is unknown. Several molecules have been identified that, like vegetal cortical cytoplasm, are able to produce a secondary axis when expressed in early embryos. Activation of the Wnt-signaling pathway by Xwnt-8 (Sokol et al., 1991; Smith and Harland, 1991), Xwnt-8b (Cui et al., 1995), Xdsh (Sokol et al., 1995), a dominant-negative form of Xgsk3 (Dominguez et al., 1995; He et al., 1995; Pierce and Kimelman, 1995), XAPC (Vlem- inckx et al., 1997), and b-catenin (Funayama et al., 1995) causes full axis formation, although none of their mrnas or proteins has been shown to be localized at the vegetal cortex of the unfertilized egg. The mrna for another Wnt family member, Xwnt-11, is localized at the oocyte vegetal cortex, but Xwnt-11 RNA is unable to produce a full dorsal axis (Ku and Melton, 1993). A secreted molecule noggin also has the ability to induce a complete dorsal axis, but noggin RNA is not localized to the vegetal cortex (Smith and Har- land, 1992). The RNA for the TGFb family member Vg1 is localized to the oocyte vegetal cortex (Weeks and Melton, /97 $25.00 Copyright 1997 by Academic Press All rights of reproduction in any form reserved. 69
2 70 Marikawa et al. 1987). Vg1 RNA has no dorsalizing activity, but BVg1, a modified version of Vg1 designed to promote efficient secretion of mature Vg1, has the ability to induce a full dorsal axis (Dale et al., 1993; Thomsen and Melton, 1993). Expression of a transcription factor Siamois is induced in animal caps by Xwnt-8, a dominant-negative form of Xgsk3, XAPC, and b-catenin, but not by BVg1 or noggin (Brannon and Kimelman, 1996; Carnac et al., 1996; Vleminckx et al., 1997). Expression of the TGFb family member Xnr3 is also induced in animal caps by Xwnt-8 but not by noggin or a dorsal mesoderm inducer, activin (Smith et al., 1995). Thus, it is likely that expression of Siamois and Xnr3 is a specific marker for activation of the Wnt pathway. In addition, b- catenin, acting as a downstream element of the Wnt pathway, translocates into the nucleus when activated (Funayama et al., 1994; Schneider et al., 1996; Yost et al., 1996; Larabell et al., 1997). Therefore, it is of particular interest to examine whether the dorsal determinants in the vegetal cortical cytoplasm are able to induce transcriptional activation of Xnr3 and Siamois and a nuclear accumulation of b- catenin protein. For this purpose, we exploited the reversed embryo, which is derived from fertilized eggs that are inverted to orient the unpigmented vegetal hemisphere upward (Neff et al., 1983). In the inverted egg, heavy yolky cytoplasm, which is originally located in the unpigmented half, is displaced into the opposite side by the gravity, while the surface pigmentation pattern is unchanged (Neff et al., 1984). The inverted egg is able to develop into an apparently normal embryo except for the pigmentation pattern. The unpigmented side gives rise to ectodermal tissues and the pigmented side gives rise to endodermal tissues (Neff et al., 1983). Thus, the embryo derived from the inverted egg displays a reversal of the animal vegetal axis with regard to the cortical pigmentation. In this study, we have determined the location of the dorsal determinants in the in- verted egg. Then, we have examined the correlation be- tween the dorsal determinants and expression pattern of Xnr3 and Siamois and the distribution of b-catenin protein in the reversed embryo. MATERIALS AND METHODS Eggs and Embryos Adult Xenopus laevis females were injected with 650 IU of human chorionic gonadotropin (Sigma, St. Louis, MO) to induce ovulation and kept at C overnight. Eggs were squeezed into sperm suspension, which was prepared by homogenizing testes in 100% Steinberg s saline. Fertilized eggs were dejellied in 2.6% L- cysteine solution (ph 8.0) and washed extensively in 20% Steinberg s saline. Inversion and Centrifugation of Eggs Neff et al. (1983, 1984) prepared inverted eggs by placing fertilized eggs upside down under natural gravity, and Black and Gerhart (1985) added a centrifugation process to promote efficient displace- FIG. 1. A schematic diagram of the procedure of inversion and centrifugation of eggs (see Materials and Methods). SS, Steinberg s saline. ment of inner cytoplasm. We have obtained inverted eggs according to the centrifugation procedure of Dr. Atsunori Shinagawa, Yamagata University, Japan (reported at the Annual Meeting of the Japanese Society of Developmental Biologists, 1995). The procedure is schematically represented in Fig. 1. Dejellied eggs were placed in 5% Ficoll dissolved in 100% Steinberg s saline to remove water in the perivitelline space. Each egg was placed in a hole made in Paraplast coating the bottom of a dish that was filled with 5% Ficoll in 100% Steinberg s saline. Eggs were inverted with the aid of a hair loop to orient the unpigmented vegetal hemisphere up- ward. At min after insemination (approximately 35% of the first cell cycle: 0.35NT), the dish was placed on the top of a centri- fuge bucket and centrifuged at 10 30g for 5 10 min. After centrifugation, eggs were transferred to 20% Steinberg s saline and cultured at 15 C. Normal looking blastulae were developed from 40 90% of inverted eggs. Cytoplasmic Transplantation Transplantation of cortical cytoplasm was carried out according to Holowacz and Elinson (1993). The tip of a glass microneedle was
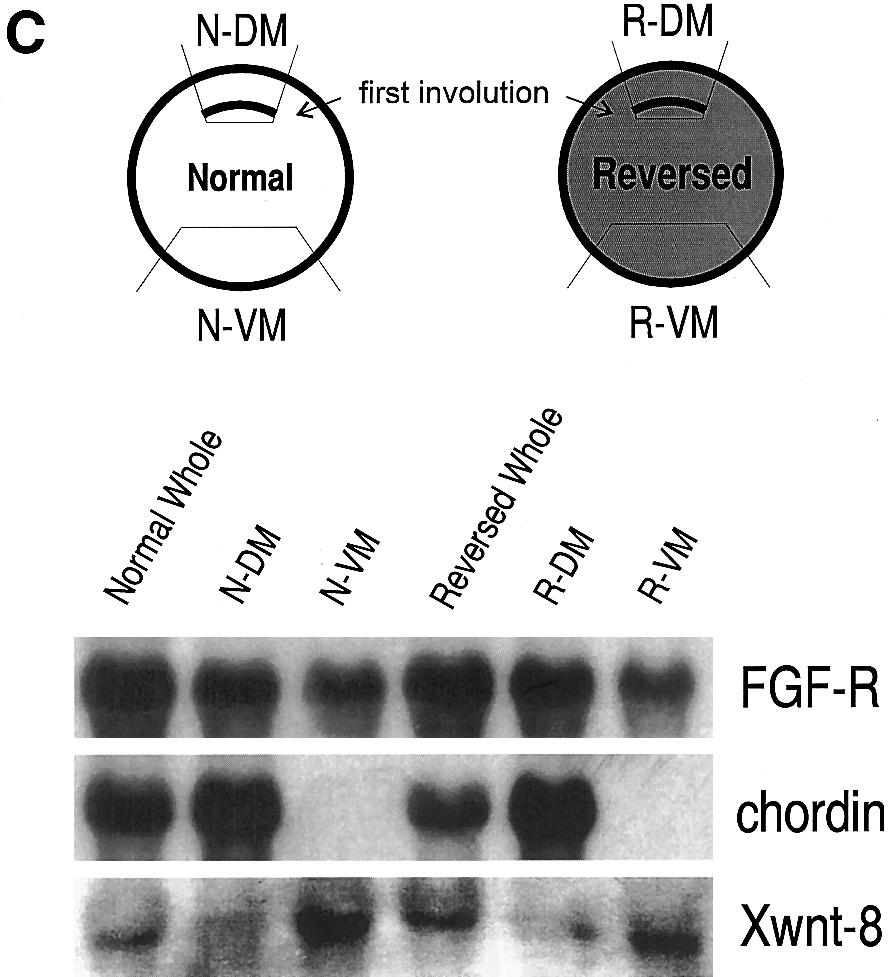
3 Xenopus Dorsal Determinants Activate Wnt Pathway 71 beveled with an inclination of 30 by a microneedle grinder (Model overnight at 020 C in 80% methanol, 20% DMSO. After several PB-7; Narishige, Tokyo). Cortical cytoplasms were withdrawn from washes in TBST (140 mm NaCl, 10 mm Tris, ph 7.4, 0.1% Triton), a normal egg and an inverted egg and injected into two ventral the embryos were blocked in 1% BSA in TBST at room temperature vegetal blastomeres of a 16-cell recipient embryo. Dorsoanterior for half an hour, followed by incubation at 4 C overnight in anti- index (DAI) of the secondary axis was scored according to Kao and b-catenin antibodies (1:100 dilution; a generous gift from Dr. Peter Elinson (1988). Hausen, Tübingen, Germany) in 1% BSA in TBST. After washing in 0.2% BSA in TBST for three times, 2 hr per wash, the embryos were incubated at 4 C overnight in AP-conjugated goat anti-rabbit Northern Analysis IgG (1:700 dilution; BIO/CAN Sci. Inc., Mississauga, Ontario) in 1% BSA in TBST. After extensive wash in 0.2% BSA in TBST, RNA was extracted from whole embryos and dissected tissue the embryos were visualized by the NBT/BCIP method (Boerhinger pieces by the Proteinase K/LiCl method according to Sambrook et Mannheim) and cleared in benzyl benzoate/benzyl alcohol. al. (1989), with a slight modification. Specimens were digested in 1 mg/ml Proteinase K, 20 mm Tris (ph 7.6), 100 mm NaCl, 30 mm EDTA, and 1% SDS at 37 C for 1 hr to overnight. After three extractions with phenol/chloroform, RNA was precipitated in 4 M RESULTS LiCl and washed in 70% ethanol. The RNA was electrophoretically separated on a 1% agarose gel containing formaldehyde and Mops buffer, blotted onto a nylon Development of the Reversed Embryo membrane (Boehringer Mannheim) by capillary transfer, and UV The centrifugally inverted eggs developed into embryos crosslinked. The membrane was prehybridized in hybridization that looked similar to those described in Neff et al. (1983). buffer (HB: 50% formamide, 51 SSC, 101 Denhardt s solution, 1 They displayed a reversal of the small/large blastomere patmg/ml torula RNA, 1% SDS, 10 mm sodium phosphate, ph 6.8) tern: smaller blastomeres developed in the unpigmented at 65 C for 1 hr and hybridized in HB containing ng/ml hemisphere and larger blastomeres developed in the pigof digoxygenin (DIG)-labeled RNA probes at 65 C overnight. DIGmented hemisphere. At the blastula stage, a blastocoel was labeled RNA probes for chordin (Sasai et al., 1994), Xwnt-8 (Chrisformed near the unpigmented hemisphere of the reversed tian et al., 1991), Vg1 (Weeks and Melton, 1987), Xcat2 (Mosquera et al., 1993), Xnr3 (Smith et al., 1995), Mix.1 (Rosa, 1989), and the embryo. At the early gastrula stage, the first involution was fibroblast growth factor receptor (FGF-R; Musci et al., 1990) were observed subequatorially as an accumulated pigmentation. made by in vitro transcription using DIG-UTP according to the The blastopore lip of the reversed embryo expanded later- manufacturer s instructions (Boehringer Mannheim). After hybridization, ally in the pigmented hemisphere (Fig. 2A). Some of the the membrane was washed at 68 C twice in 21 SSC, 0.1% reversed embryos (20 90% of normal looking blastulae deally SDS and twice in 0.11 SSC, 0.1% SDS, 15 min per wash. The pending on the batches of the eggs used) developed into membrane was then rinsed in maleic acid buffer (MAB: 0.1 M ma- apparently normal tadpoles except for the pigmentation leic acid, 0.15 M NaCl, ph 7.5), blocked in MAB containing 1% pattern: the anterior portion of the reversed embryo was blocking reagent (Boehringer Mannheim), and incubated in MAB less pigmented compare to that of the normal tadpole (Fig. containing 1% blocking reagent plus alkaline phosphatase (AP)- 2C). In most cases, the cement gland of the reversed embryo conjugated anti-dig antibody (1:5000 dilution; Boehringer Mannwas totally unpigmented. Therefore, as reported by Neff et heim). The membrane was then washed twice in MAB containing 0.15% Tween 20, 20 min per wash, and rinsed in alkaline phosphaanimal vegetal axis with regard to the cortical pigmenta- al. (1983), the inversion of eggs resulted in the reversal of tase buffer (APB: 0.1 M Tris HCl, 0.1 M NaCl, ph 9.5). After incution bation in APB containing chemiluminescent substrate CSPD (1:100 pattern. dilution; Boehringer Mannheim), the membrane was exposed to an To see whether the site of the first involution of the X-ray film. reversed embryo corresponds to a dorsal lip, the expression pattern of chordin, whose transcript is found exclusively RT PCR Analysis on the dorsal lip in the normal embryo (Sasai et al., 1994), was examined. Chordin transcript was present at the site RNA was isolated from whole embryos or animal caps using of first involution but not on the side opposite to the first Trizol reagent (Gibco BRL, Gaithersburg, MD) following the manu- involution (Fig. 2C). In agreement, when the expression of facturer s instructions. DNA was removed by digestion with RQ1 the ventral lateral mesoderm marker Xwnt-8 (Christian et DNase (Promega, Madison, WI) at 37 C for half an hour. Reverse al., 1991) was examined, it was found more strongly on the transcription and PCR conditions were according to Hemmati-Bri- side opposite to the first involution (Fig. 2C). Thus, the vanlou and Melton (1994). EF-1a primers were as described by Hemfirst involution of the reversed embryo corresponds to a mati-brivanlou and Melton (1994). Siamois primers were as dedorsal lip. scribed by Brannon and Kimelman (1996). PCR products were analyzed on a 5% acrylamide gel. Association of Dorsal Determinants with the Immunocytochemistry Cortical Cytoplasm of the Unpigmented Hemisphere Whole mount immunostaining for b-catenin was carried out according to Schneider et al. (1996) with the following modification. Dorsal determinants are present in the vegetal cortical Embryos were fixed for 2 3 hr with 3.7% formaldehyde in 0.1 cytoplasm of the normal egg (Fujisue et al., 1993; Holowacz M Mops (ph 7.4), 2 mm EGTA, 1 mm MgSO 4 and permeabilized and Elinson, 1993; Kikkawa et al., 1996; Sakai, 1996). To

4 72 Marikawa et al.
5 Xenopus Dorsal Determinants Activate Wnt Pathway 73 nal mrnas, Vg1 and Xcat2. Vg1 RNA is localized at the vegetal cortex of oocytes. The RNA is released from its tight localization after maturation but remains in the vegetal hemisphere (Melton, 1987; Weeks and Melton, 1987). Xcat2 RNA is associated with the vegetal cortex and remains associated with the cortex after maturation (Mosquera et al., 1993; Forristall et al., 1995). To examine the distribution of Vg1 and Xcat2 RNAs in the inverted egg, cytoplasmic fragments (each approximately one-fifth of total egg volume) were cut from the reversed animal and the reversed vegetal poles and analyzed by Northern blotting (Fig. 4A). A significant amount of Vg1 RNA was found in the reversed animal fifth as well as in the reversed vegetal fifth (Fig. 4B), suggesting that a certain amount of Vg1 RNA may be still associated with the cortical cytoplasm even after the release from its tight localization during oocyte maturation. Xcat2 RNA was also found in the reversed animal fifth. The amount of Xcat2 RNA in the reversed animal fifth was almost as much as in the normal vegetal fifth (Figs. 4C and 4D). However, the presence or absence of Xcat2 RNA in the reversed vegetal fifth was variable. In one case, less Xcat2 RNA was found in the reversed vegetal fifth (Fig. 4D). But in another case, Xcat2 RNA was not detected in the reversed vegetal fifth (Fig. 4C). A similar difference in Xcat2 RNA distribution was observed when the reversed animal and the reversed vegetal halves were examined: the RNA was exclusively found in the reversed animal half in one case, but the RNA was also found in the reversed vegetal half in the other case (data not shown). The reason for this difference is not clear, although variation in cytoplasmic fluidity among batches of eggs may affect how molecules are redistributed by centrifugation (Neff et al., 1990). Alter- natively, there may be a certain variation in the RNA binding capability of the cortex depending on the batches of eggs. In any case, the reversed animal cortex of the inverted egg contained a significant amount of Vg1 and Xcat2 RNAs. examine the location of the dorsal determinants in the inverted egg, cytoplasmic transplantation was performed. To avoid confusion, parts of the reversed egg are named as shown in Fig. 1. The unpigmented hemisphere of the reversed egg is called the reversed animal hemisphere because it is fated to become ectoderm, and the pigmented hemisphere of the reversed egg is called the reversed vegetal hemisphere because it is fated to become endoderm. The animal and vegetal hemispheres of the normal egg are called the normal animal and normal vegetal hemispheres, respectively. In both the normal and the reversed cases, the vegetal hemisphere contains the mass of large yolk platelets (Fig. 1). Cortical cytoplasms were injected into two ventral vegetal blastomeres of a normal 16-cell recipient embryo and assayed for the ability to induce a secondary axis (Fig. 3). The cortical cytoplasm of the normal vegetal hemisphere induced a secondary axis (6/13). In contrast, the cortical cytoplasm of the reversed vegetal hemisphere did not induce a secondary axis (0/14), while the reversed animal cortical cytoplasm induced an axis (12/16). The axis forming activity of the reversed animal cortical cytoplasm (average DAI Å 2.1) was as strong as the normal vegetal cortical cytoplasm (average DAI Å 1.4). The presence of dorsal determinants in the animal cortical cytoplasm of the reversed egg may indicate that the dorsal determinants are associated with the unpigmented cortex firmly enough to resist the centrifugal force. Alternatively, the dorsal determinants may be associated with cytoplasmic components that had low specific gravity and end up in the centripetal (reversed animal) side of centrifuged eggs. In order to discriminate these possibilities, eggs were centrifuged in the uninverted orientation (Fig. 3C, bottom row). The cortical cytoplasm of the pigmented hemisphere, located at the centripetal side of these centrifuged eggs, did not induce a secondary axis (0/9 cases). However, the cortical cytoplasm of the unpigmented hemisphere of these centrifuged eggs induced a secondary axis (8/12; average DAI Å 1.1). These results indicate that the dorsal determinants are firmly associated with the unpigmented cortex. Distribution of Localized Maternal Messenger RNAs in the Inverted Egg Because the dorsal determinant activity was found exclusively in the reversed animal cortex, it is of particular interest to see what kind of molecules are associated with the reversed animal cortex. We focused on two localized mater- Expression of Xnr3 and Siamois in Ectodermal Cells of the Reversed Embryo The TGFb family member Xnr3 is expressed in the superficial layer of Spemann organizer (Smith et al., 1995), and the homeobox-containing transcription factor Siamois is expressed within the Nieuwkoop Center (Lemaire et al., 1995). Xnr3 and Siamois are expressed in animal cap cells in response to several dorsalizing genes which are components and activators of the Wnt pathway, but they are not induced by noggin or dorsal mesoderm inducers including FIG. 2. Development of the reversed embryos. (A) A vegetal view of a normal (N) and a reversed (R) embryo at gastrula stage. An expanding blastopore lip is seen in the pigmented area of the reversed embryo. (B) A normal (N) and a reversed (R) embryo at late tailbud stage. Note that the anterior portion, especially the cement gland, of the reversed embryo is much less pigmented than that of the normal embryo. (C) Expression pattern of Chordin and Xwnt-8 in the reversed embryo. Marginal tissues were taken from early gastrulae as shown in the diagram and analyzed by Northern blot. Chordin RNA is expressed exclusively at the site of the first involution (N-DM and R- DM), while Xwnt-8 RNA is expressed more on the side opposite to the first involution (N-VM and R-VM). FGF-R is used as a loading control.
 A schematic diagram showing the experimental procedure used to assay for the ability of cortical cytoplasm to induce a secondary axis.")
 Examples of double axis embryos that were produced by injection of the cortical cytoplasm from the reversed animal hemisphere of the inverted egg.")
6 74 Marikawa et al. FIG. 3. Dorsalizing activity of cortical cytoplasm. (A) A schematic diagram showing the experimental procedure used to assay for the ability of cortical cytoplasm to induce a secondary axis. Cortical cytoplasms from the animal pole or the vegetal pole were withdrawn from donor eggs and microinjected into two ventral vegetal blastomeres of a 16-cell recipient embryo. (B) Examples of double axis embryos that were produced by injection of the cortical cytoplasm from the reversed animal hemisphere of the inverted egg. (C) A table summarizing the result of the cytoplasmic transfer experiment. Top row: Cortical cytoplasm taken from the vegetal hemisphere of the normal egg had dorsal axis forming activity. Middle row: Cortical cytoplasms of the inverted egg were taken from the unpigmented (reversed animal) and the pigmented (reversed vegetal) hemispheres. Dorsal axis forming activity was found mostly in the reversed animal cortical cytoplasm. Bottom row: Cortical cytoplasms were taken from the pigmented and the unpigmented hemispheres of the egg that is centrifuged without inversion. Dorsal axis forming activity was found exclusively in the unpigmented cortical cytoplasm. Dorsoanterior index (DAI) of the secondary axis was scored according to Kao and Elinson (1988). that the agent functions similarly to the activators of the Wnt pathway. In the inverted eggs, the dorsal determinants were located in the reversed animal hemisphere, which was fated to be- BVg1 and activin (Smith et al., 1995; Carnac et al., 1996; Brannon and Kimelman, 1996; Fagotto et al., 1997). Therefore, if an unidentified dorsalizing agent induces the expression of Xnr3 and Siamois in animal cap cells, it is likely
 Cytoplasmic fragments (each approximately one-fifth of total egg volume) were dissected from eggs that were fixed in 98% ethanol and 2% acetic acid and digested in the Proteinase K solution for")
7 Xenopus Dorsal Determinants Activate Wnt Pathway 75 FIG. 4. Distribution of the Xcat2 and Vg1 RNAs in the reversed egg. (A) Cytoplasmic fragments (each approximately one-fifth of total egg volume) were dissected from eggs that were fixed in 98% ethanol and 2% acetic acid and digested in the Proteinase K solution for RNA extraction. (B) A Northern blot of Vg1. The RNA is present in the reversed animal fifth as well as the reversed vegetal fifth. (C) A Northern blot of Xcat2. The RNA is seen exclusively in the reversed animal fifth. (D) Another Northern blot of Xcat2. The RNA is present more in the reversed animal fifth and less in the reversed vegetal fifth. Each lane contains total RNA extracted from five cytoplasmic fragments. FGF-R was used as a loading control. come ectoderm. This situation offers an opportunity to examine whether the dorsal determinants in ectodermal cells induce Xnr3 and Siamois. Animal caps were taken from reversed embryos at the early gastrula stage and assayed for the expression of these genes (Fig. 5A). Both Xnr3 and Siamois were expressed in the reversed animal cap, but neither of them was expressed in the normal animal cap (Figs. 5B and 5C), suggesting that the dorsal determinants behave like activators of the Wnt pathway. Mix.1 is expressed in endodermal and mesodermal cells in normal development and is induced in animal caps by dorsal mesoderm inducers including BVg1 and activin (Rosa, 1989; Cornell et al., 1995). Mix.1 was strongly expressed in the reversed vegetal cap but not expressed in the reversed animal caps (Fig. 5D). Therefore, the dorsal determinants are unlikely to be dorsal mesoderm inducers like BVg1 and activin, consistent with the previous study (Holowacz and Elinson, 1995). Furthermore, the Mix.1 expression pattern confirmed a reversal of the animal vegetal axis in the reversed embryo with respect to the endoderm. Accumulation of b-catenin Protein in Nuclei of Animal Pole Cells of the Reversed Embryo Schneider et al. (1996) and Larabell et al. (1997) have shown that b-catenin protein, acting as a component of the Wnt-signaling pathway, accumulates in nuclei on the dorsal side of the blastula. Several treatments which result in Wnt- pathway activation, such as mrna injections of Xwnt-8, a dominant-negative mutant of Xgsk3, and b-catenin, induce a nuclear accumulation of b-catenin protein (Funayama et al., 1995; Schneider et al., 1996; Yost et al., 1996; Larabell et al., 1997). Since the dorsal determinants behave like the Wnt-pathway activators with regard to the transcriptional activation of Xnr3 and Siamois in the animal cap, it is of interest to examine whether b-catenin protein accumulates in nuclei of cells in the reversed animal hemisphere. At stage 8 9, whole reversed and normal embryos were immunocytochemically stained with anti-b-catenin polyclonal antibody. b-catenin was found along the plasma membrane of all cells of both the reversed and the normal embryos, probably due to the cadherin/catenin interaction. In 23 of 58 reversed embryos from four experiments, a patch of cells near the animal pole showed strong nuclear staining, while surrounding animal cells did not (Fig. 6A). Cells with nuclear staining often also had some cytoplasmic staining. An additional 14 reversed embryos had nuclear staining in animal pole cells, and the nuclear staining extended down into the marginal zone. In contrast, animal cells of normal embryos did not show strong nuclear staining, although weak dumbbell-shaped staining was observed in most cells (Fig. 6B). The latter staining was probably perinuclear, as noted by Schneider et al. (1996). Nuclei were stained on one side of the marginal zone of the normal embryo (25 out of 47), confirming that our immunostaining procedure was appropriate (Fig. 6C). The observation of nuclear staining
 Animal caps and vegetal caps were taken from the normal embryo and the reversed embryo at early gastrulae stage (stage 10/).")
 An RT PCR showing the expression pattern of Siamois.")
8 76 Marikawa et al. FIG. 5. Expression pattern of Xnr3, Siamois, and Mix.1 in the reversed embryos. (A) Animal caps and vegetal caps were taken from the normal embryo and the reversed embryo at early gastrulae stage (stage 10/). (B) A Northern blot showing the expression pattern of Xnr3, in a stage 10/ normal whole embryo, five normal animal caps, and five reversed animal caps. Xnr3 is strongly expressed in the reversed animal cap but not expressed in the normal animal cap. FGF-R was used as a loading control. (C) An RT PCR showing the expression pattern of Siamois. Siamois is expressed in the reversed animal cap but not expressed in the normal animal cap. EF-1a was used as a control. (D) A Northern blot showing the expression pattern of Mix.1. Each lane contains total RNA corresponding to a single whole embryo or five caps. Mix.1 is expressed in both the normal vegetal cap and the reversed vegetal cap, but in neither the normal animal cap nor the reversed animal cap. FGF-R was used as a loading control. in about half of both reversed and normal embryos is in part due to the lumping of data from stage 8 through stage 9 embryos. The older embryos with smaller cells tended not to have nuclear staining. DISCUSSION The egg vegetal cortex contains the dorsal determinants that are responsible for establishing dorsoventral patterning FIG. 6. Nuclear accumulation of b-catenin protein in the animal hemisphere of the reversed embryo. (A) A reversed embryo (stage 8.5) had a patch of cells near the animal pole in which nuclei are stained strongly compare to the surrounding cells. The dark area at the bottom is displaced animal pigment, much of which is visible because the embryo is cleared. (B) A normal albino embryo (stage 8.5) which was injected with the mrna for a dominant negative mutant of XGSK3 into one animal blastomere at the 8-cell stage as a positive control (Dominguez et al., 1995; Yost et al., 1996). A patch of cells in the animal hemisphere had strong nuclear staining (arrow), while the surrounding animal region did not. (C) A normal albino embryo (stage 8.5) had strong nuclear staining on one side of the marginal zone.
9 Xenopus Dorsal Determinants Activate Wnt Pathway 77 direct synthesis of a processed mature form of Vg1 and induce axial structures in injected embryos (Dale et al., 1993; Thomsen and Melton, 1993). It may be that endogenous Vg1 precursor protein is processed specifically on the dorsal side of normal embryos, thus allowing Vg1 to act in setting up the dorsal axis. However, in the reversed egg, a signifi- cant amount of Vg1 mrna is present at the animal pole, which contains the dorsal axis forming activity. Nonethe- less, Mix.1, which is induced in normal animal caps by BVg1 (Cornell et al., 1995), is expressed at the vegetal pole but not at the animal pole of the reversed embryo. This implies that Vg1 is not processed in the animal region of the reversed embryo, even though the dorsal determinants are present in the same place. The dorsal determinants are located in the future ectodermal region in the inverted egg, raising the question how the reversed embryo gives rise to a normal tadpole. In normal development, the dorsal determinants are thought to estab- lish the Nieuwkoop Center, which is located at the dorsal vegetal region and has endodermal fate. The Nieuwkoop Center induces the Spemann organizer in the overlying marginal region in turn. We do not know whether the ectoder- mal tissue, which has received the dorsal determinants in the reversed embryo, behaves like the Nieuwkoop Center to induce Spemann organizer in the underlying marginal region. Wylie et al. (1996) has shown that a dorsal axis in the b-catenin-depleted embryo is recovered by injection of b-catenin mrna into the most animally located blasto- mere, even though the injected blastomere gives rise to the anterior epidermis. In the present study, we showed that the dorsal determinants in animal cap cells induce expression of two Wnt-target genes, Xnr3 and Siamois, and nuclear accumulation of b-catenin. Therefore, dorsal determinants located at the animal pole in the reversed embryo may direct formation of the Spemann organizer in the underlying marginal region by a mechanism similar to the rescue of the embryonic axis with animal b-catenin injections. Alternatively, although we could only detect dorsal determinants in the reversed animal hemisphere by cytoplasmic transplantation, the dorsal determinants may spread into the marginal zone, as suggested by our observation that some reversed embryos had b-catenin nuclear staining at the mar- ginal zone as well as at the animal pole. Further experiments, such as deletions of regions of the inverted egg, are required to determine where the active dorsal center is in the reversed embryo. of the Xenopus embryo. To understand the molecular nature of the dorsal determinants, we exploited the animal vegetal reversed embryo that is derived from the inverted egg. The dorsal determinants in the inverted egg were located at the unpigmented cortex, which was segregated into ectodermal tissues. Animal cap cells of the reversed embryos revealed the expression of Xnr3 and Siamois, as well as the nuclear accumulation of b-catenin, while they did not express Mix.1. Our results on the endogenous dorsal determinant are concordant with a variety of results in which members of the Wnt/b-catenin pathway are overexpressed or underexpressed (Heasman et al., 1994; Wylie et al., 1996; Fagotto et al., 1996, 1997). Therefore, we conclude that the dorsal determinants of the egg behave more like the Wnt-pathway activators rather than noggin or BVg1. The reversed embryo provides a direct correlation between the dorsal determinants of the vegetal cortical cytoplasm, the transcriptional activation of Xnr3 and Siamois, and the nuclear accumulation of b-catenin protein. A similar correlation between the dorsal determinants and activation of the Wnt pathway is seen in embryos that are derived from UV-treated eggs. The UV-treated egg retains the dorsal axis forming activity at the vegetal pole, as demonstrated by cytoplasmic transfer (Fujisue et al., 1993; Holowacz and Elinson, 1993). Vegetal pole cells of the UV-treated embryo also express Siamois (Brannon and Kimelman, 1996; Cui et al., 1996) and have b-catenin protein translocated into nuclei (Schneider et al., 1996). Therefore, the dorsal determinants may activate the Wnt-signaling cascade regardless of their location in the embryo. Our cytoplasmic transfer results indicate that the cortical dorsal activity remained with the unpigmented cortex in the reversed embryos. This result suggests that the dorsal determinants are firmly associated with the vegetal cortex and allows us to estimate their distance from the surface. Work by others has shown that after inversion of the egg, the germ plasm remained localized at the unpigmented surface and is partitioned into reversed animal cap cells. The reversed embryos contained a reduced number of the primordial germ cells (PGCs), probably as a result of the displacement of the germ plasm away from its original vegetal pole location (Neff et al., 1983; Wakahara et al., 1984; Cleine and Dixon, 1985; Cleine, 1986; Jurand and Dixon, 1986). Xcat2 mrna appears to be associated with the germ plasm (Forristall et al., 1995; Kloc and Etkin, 1995). In our inverted eggs, Xcat2 mrna stayed mostly at the reversed animal pole but some did not, indicating that the germ plasm tended to remain with the unpigmented surface. Since the germ plasm is distributed within mm from the egg surface (Ressom and Dixon, 1988), the bulk of the cortical dorsal determinants must be distributed within this distance from the egg surface. Kageura s (1997) recent dem- onstration of dorsal determinants in isolated cortex also indicates that the determinants are localized very close to the surface. Injection of Vg1 mrna directs synthesis of Vg1 precursor protein but not a mature product, resulting in no effect on normal development. In contrast, the BVg1 mrna can ACKNOWLEDGMENTS We thank A. Shinagawa, who presented experiments on the centrifugally reversed embryo at the Annual Meeting of the Japanese Society of Developmental Biologists (1995), for inspiring Y.M. We are grateful for the invaluable gift of the anti-b-catenin antibody from P. Hausen. We also thank M. Brannon and D. Kimelman for their technical advice on the RT PCR conditions, E. Amaya for FGF-R plasmid, E. M. DeRobertis for chordin plasmid, D. Kimelman for Mix.1 plasmid, M. L. King for Xcat2 plasmid, D. A. Melton
10 78 Marikawa et al. for Vg1 plasmid, S. Sokol for Xwnt-8 and a dominant negative mu- Funayama, N., Fagotto, F., McCrea, P., and Gumbiner, B. M. (1995). tant of XGSK3 plasmids, and W. C. Smith and R. M. Harland for Embryonic axis induction by the armadillo repeat domain of b- Xnr3 plasmid. We acknowledge Y. Masui and members of the Elin- catenin: Evidence for intracellular signaling. J. Cell. Biol. 128, son lab for their helpful discussion. Y.M. is supported by a postdoctoral fellowship from the Human Frontier Science Program Organi- He, X., Saint-Jeannet, J-P., Woodgett, J. R., Varmus, H. E., and zation. This work was funded by a grant to R.P.E. from Medical Dawid, I. B. (1995). Glycogen synthase kinase-3 and dorsoventral Research Council of Canada. patterning in Xenopus embryos. Nature 374, Heasman, J., Crawford, A., Goldstone, K., Garner-Hamrick, P., Gumbiner, B., McCrea, P., Kintner, C., Yoshida-Noro, C., and Wylie, C. (1994). Overexpression of cadherins and underexpres- REFERENCES sion of b-catenin inhibit dorsal mesoderm induction in early Xenopus embryos. Cell 79, Black, S. D., and Gerhart, J. C. (1985). Experimental control of the Hemmati-Brivanlou, A., and Melton, D. A. (1994). Inhibition of site of embryonic axis formation in Xenopus laevis eggs centri- activin receptor signaling promotes neuralization in Xenopus. fuged before first cleavage. Dev. Biol. 108, Cell 77, Brannon, M., and Kimelman, D. (1996). Activation of Siamois by Holowacz, T., and Elinson, R. P. (1993). Cortical cytoplasm, which the Wnt pathway. Dev. Biol. 180, induces dorsal axis formation in Xenopus, is inactivated by UV Carnac, G., Kodjabachian, L., Gurdon, J. B., and Lemaire, P. (1996). irradiation of the oocyte. Development 119, The homeobox gene Siamois is a target of the Wnt dorsalisation Holowacz, T., and Elinson, R. P. (1995). Properties of the dorsal pathway and triggers organiser activity in the absence of meso- activity found in the vegetal cortical cytoplasm of Xenopus eggs. derm. Development 122, Development 121, Christian, J. L., McMahon, J. L., McMahon, A. P., and Moon, R. T. Jurand, A., and Dixon, K. E. (1986). The fine structure of germ plasm (1991). Xwnt-8, a Xenopus Wnt-1/int-related gene responsive to in eight-cell embryos developing from normal and inverted eggs mesoderm-inducing growth factors, may play a role in ventral of Xenopus laevis. J. Exp. Zool. 237, mesodermal patterning during embryogenesis. Development Kageura, H. (1997). Activation of dorsal development by contact 111, between the cortical dorsal determinant and the equatorial core Cleine, J. H. (1986). Replacement of posterior by anterior endoderm cytoplasm in eggs of Xenopus laevis. Development 124, 1543 reduces sterility in embryos from inverted eggs of Xenopus laevis J. Embryol. Exp. Morphol. 94, Kao, K. R., and Elinson, R. P. (1988). The entire mesodermal mantle Cleine, J. H., and Dixon, K. E. (1985). The effect of egg rotation on behaves as Spemann s organizer in dorsoanterior enhanced Xenothe differentiation of primordial germ cells in Xenopus laevis. J. pus laevis embryos. Dev. Biol. 127, Embryol. Exp. Morphol. 90, Kikkawa, M., Takano, K., and Shinagawa, A. (1996). Localization Cornell, R. A., Musci, T. J., and Kimelman, D. (1995). FGF is a and behavior of dorsal determinants during first cell cycle in prospective competence factor for early activin-type signals in Xenopus eggs. Development 122, Xenopus mesoderm induction. Development 121, Kloc, M., and Etkin, L. D. (1995). Two distinct pathways for the Cui, Y., Brown, J. D., Moon, R. T., and Christian, J. L. (1995). Xwntlocalization of RNAs at the vegetal cortex in Xenopus oocytes. 8b: A maternally expressed Xenopus Wnt gene with a potential Development 121, role in establishing the dorsoventral axis. Development 121, Ku, M., and Melton, D. A. (1993). Xwnt-11: A maternally expressed Cui, Y., Tian, Q., and Christian, J. L. (1996). Synergistic effects of Xenopus wnt gene. Development 119, Vg1 and Wnt signals in the specification of dorsal mesoderm and Larabell, C. A., Torres, M., Rowning, B. A., Yost, C., Miller, J. R., endoderm. Dev. Biol. 180, Wu, M., Kimelman, D., and Moon, R. T. (1997). Establishment Dale, L., Matthew, G., and Colman, A. (1993). Secretion and mesoasymmetries in beta-catenin that are modulated by the Wnt sig- of the dorso-ventral axis in Xenopus embryos is presaged by early derm-inducing activity of the TGF-b-related domain of Xenopus Vg1. EMBO J. 12, naling pathway. J. Cell Biol. 136, Dominguez, I., Itoh, K., and Sokol, S. Y. (1995). Role of glycogen Lemaire, P., Garrett, N., and Gurdon, J. B. (1995). Expression clonsynthase kinase 3b as a negative regulator of dorsoventral axis ing of Siamois, a Xenopus homeobox gene expressed in dorsal- formation in Xenopus embryos. Proc. Natl. Acad. Sci. USA 92, vegetal cells of blastulae and able to induce a complete secondary axis. Cell 81, Fagotto, F., Funayama, N., Glück, U., and Gumbiner, B. M. (1996). Melton, D. A. (1987). Translocation of a localized maternal mrna Binding to cadherins antagonizes the signaling activity of b-ca- to the vegetal pole of Xenopus oocytes. Nature 328, tenin during axis formation of Xenopus. J. Cell Biol. 132, 1105 Mosquera, L., Forristall, C., Zhou, Y., and King, M. L. (1993). A mrna localized to the vegetal cortex of Xenopus oocytes en- Fagotto, F., Guger, K., and Gumbiner, B. M. (1997). Induction of codes a protein with a nanos-like zinc finger domain. Developthe primary dorsalizing center in Xenopus by the Wnt/GSK/b- ment 117, catenin signaling pathway, but not by Vg1, Activin or Noggin. Musci, T. J., Amaya, E., and Kirschner, M. W. (1990). Regulation of Development 124, the fibroblast growth factor receptor in early Xenopus embryos. Forristall, C., Pondel, M., Chen, L., and King, M. L. (1995). Patterns Proc. Natl. Acad. Sci. USA 87, of localization and cytoskeletal association of two vegetally lo- Neff, A. W., Malacinski, G. M., Wakahara, M., and Jurand, A. calized RNAs, Vg1 and Xcat-2. Development 121, (1983). Pattern formation in amphibian embryos prevented from Fujisue, M., Kobayakawa, Y., and Yamana, K. (1993). Occurrence undergoing the classical rotation response to egg activation. of dorsal axis-inducing activity around the vegetal pole of an Dev. Biol. 97, uncleaved Xenopus egg and displacement to the equatorial region Neff, A. W., Wakahara, M., Jurand, A., and Malacinski, G. M. by cortical rotation. Development 118, (1984). Experimental analyses of cytoplasmic rearrangements
11 Xenopus Dorsal Determinants Activate Wnt Pathway 79 which follow fertilization and accompany symmetrization of in- Smith, W. C., McKendry, R., Ribisi, S., and Harland, R. M. (1995). verted Xenopus eggs. J. Embryol. Exp. Morphol. 80, A nodal-related gene defines a physical and functional domain Neff, A. W., Wakahara, M., and Malacinski, G. M. (1990). Bifurca- within the Spemann organizer. Cell 82, tion of the amphibian embryo s axis: Analysis of variation in Sokol, S., Christian, J. L., Moon, R. T., and Melton, D. A. (1991). response to egg centrifugation. Int. J. Dev. Biol. 34, Injected Wnt RNA induces a complete body axis in Xenopus Pierce, S. B., and Kimelman, D. (1995). Regulation of Spemann orga- embryos. Cell 67, nizer formation by the intracellular kinase Xgsk-3. Development Sokol, S., Klingensmith, J., Perrimon, N., and Itoh, K. (1995). Dor- 121, salizing and neuralizing properties of Xdsh, a maternally ex- Ressom, R. E., and Dixon, K. E. (1988). Relocation and reorganiza- pressed Xenopus homolog of dishevelled. Development 121, tion of germ plasm in Xenopus embryos after fertilization. Devel opment 103, Thomsen, G. H., and Melton, D. A. (1993). Processed Vg1 protein Rosa, F. M. (1989). Mix.1, a homeobox RNA inducible by mesoderm is an axial mesoderm inducer in Xenopus. Cell 74, inducers, is expressed mostly in the presumptive endodermal Vleminckx, K., Wong, E., Guger, K., Rubinfeld, B., Polakis, P., and cells of Xenopus embryos. Cell 57, Gumbiner, B. M. (1997). Adenomatous polyposis coli tumor sup- Sakai, M. (1996). The vegetal determinants required for the Spemresulting pressor protein has signaling activity in Xenopus laevis embryos ann organizer move equatorially during the first cell cycle. Development in the induction of an ectopic dorsoanterior axis. J. Cell 122, Biol. 136, Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular Wakahara, M., Neff, A. W., and Malacinski, G. M. (1984). Topology Cloning: A Laboratory Manual, 2nd ed. Cold Spring Harbor Labinverted of the germ plasm and development of primordial germ cells in oratory Press, Cold Spring Harbor, New York. amphibian eggs. Differentiation 26, Sasai, Y., Lu, B., Steinbeisser, H., Geissert, D., Gont, L. K., and Weeks, D. L., and Melton, D. A. (1987). A maternal mrna local- De Robertis, E. M. (1994). Xenopus chordin: A novel dorsalizing ized to the vegetal hemisphere in Xenopus eggs codes for a growth factor activated by organizer-specific homeobox genes. Cell 79, factor related to TGF-b. Cell 51, Wylie, C., Kofron, M., Payne, C., Anderson, R., Hosobuchi, M., Schneider, S., Steinbeisser, H., Warga, R. M., Hausen, P. (1996). Joseph, E., and Heasman, J. (1996). Maternal b-catenin establishes b-catenin translocation into nuclei demarcates the dorsalizing a dorsal signal in early Xenopus embryos. Development 122, centers in frog and fish embryos. Mech. Dev. 57, Smith, W. C., and Harland, R. M. (1991). Injected Xwnt-8 RNA acts Yost, C., Torres, M., Miller, J. R., Huang, E., Kimelman, D., and early in Xenopus embryos to promote formation of a vegetal Moon, R. T. (1996). The axis-inducing activity, stability, and subdorsalizing center. Cell 67, cellular distribution of b-catenin is regulated in Xenopus em- Smith, W. C., and Harland, R. M. (1992). Expression cloning of noggin, bryos by glycogen synthase kinase 3. Genes Dev. 10, a new dorsalizing factor localized to the Spemann organizer Received for publication May 21, 1997 in Xenopus embryos. Cell 70, Accepted August 11, 1997
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018
 Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Mesoderm Induction CBT, 2018 Hand-out CBT March 2018 Introduction 3. Books This module is based on the following books: - 'Principles of Developement', Lewis Wolpert, et al., fifth edition, 2015 - 'Developmental
Maternal Control of GermLayer Formation in Xenopus
 Maternal Control of GermLayer Formation in Xenopus The zygotic genome is activated at the mid-blastula transition mid-blastula fertilized egg Xenopus gastrulae early-gastrula 7 hrs 10 hrs control not VP
Maternal Control of GermLayer Formation in Xenopus The zygotic genome is activated at the mid-blastula transition mid-blastula fertilized egg Xenopus gastrulae early-gastrula 7 hrs 10 hrs control not VP
Maternal VegT and b-catenin: Patterning the Xenopus Blastula
 CHAPTER 1 Maternal VegT and b-catenin: Patterning the Xenopus Blastula Matthew Kofron, Jennifer Xanthos, and Janet Heasman 1 1.1 Introduction Loss of the maternal T-box transcription factor VegT has a
CHAPTER 1 Maternal VegT and b-catenin: Patterning the Xenopus Blastula Matthew Kofron, Jennifer Xanthos, and Janet Heasman 1 1.1 Introduction Loss of the maternal T-box transcription factor VegT has a
FORMATION AND FUNCTION OF SPEMANN S ORGANIZER
 Annu. Rev. Cell Dev. Biol. 1997. 13:611 67 Copyright c 1997 by Annual Reviews Inc. All rights reserved FORMATION AND FUNCTION OF SPEMANN S ORGANIZER Richard Harland and John Gerhart Department of Molecular
Annu. Rev. Cell Dev. Biol. 1997. 13:611 67 Copyright c 1997 by Annual Reviews Inc. All rights reserved FORMATION AND FUNCTION OF SPEMANN S ORGANIZER Richard Harland and John Gerhart Department of Molecular
Ventral mesoderm induction and patterning by bone morphogenetic protein heterodimers in Xenopus embryos
 Mechanisms of Development 74 (1998) 75 88 Ventral mesoderm induction and patterning by bone morphogenetic protein heterodimers in Xenopus embryos Shin-ichiro Nishimatsu, Gerald H. Thomsen* Department of
Mechanisms of Development 74 (1998) 75 88 Ventral mesoderm induction and patterning by bone morphogenetic protein heterodimers in Xenopus embryos Shin-ichiro Nishimatsu, Gerald H. Thomsen* Department of
Patterning the Xenopus blastula
 Development 124, 4179-4191 (1997) Printed in Great Britain The Company of Biologists Limited 1997 DEV7602 Review Article 4179 Patterning the Xenopus blastula Janet Heasman* Institute of Human Genetics
Development 124, 4179-4191 (1997) Printed in Great Britain The Company of Biologists Limited 1997 DEV7602 Review Article 4179 Patterning the Xenopus blastula Janet Heasman* Institute of Human Genetics
Expression Cloning of noggin, a New Dorsalizing Factor Localized to the Spemann Organizer in Xenopus Embryos
 Cell, Vol. 70, 829-840. September 4, 1992, Copyright 0 1992 by Cell Press Expression Cloning of noggin, a New Dorsalizing Factor Localized to the Spemann Organizer in Xenopus Embryos William C. Smith and
Cell, Vol. 70, 829-840. September 4, 1992, Copyright 0 1992 by Cell Press Expression Cloning of noggin, a New Dorsalizing Factor Localized to the Spemann Organizer in Xenopus Embryos William C. Smith and
1. What are the three general areas of the developing vertebrate limb? 2. What embryonic regions contribute to the developing limb bud?
 Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Study Questions - Lecture 17 & 18 1. What are the three general areas of the developing vertebrate limb? The three general areas of the developing vertebrate limb are the proximal stylopod, zeugopod, and
Alison Krufka,* Ross G. Johnson,*,1 Chris C. Wylie,, and Janet Heasman,, INTRODUCTION
 DEVELOPMENTAL BIOLOGY 200, 92 102 (1998) ARTICLE NO. DB988951 Evidence That Dorsal Ventral Differences in Gap Junctional Communication in the Early Xenopus Embryo Are Generated by -Catenin Independent
DEVELOPMENTAL BIOLOGY 200, 92 102 (1998) ARTICLE NO. DB988951 Evidence That Dorsal Ventral Differences in Gap Junctional Communication in the Early Xenopus Embryo Are Generated by -Catenin Independent
Early Development in Invertebrates
 Developmental Biology Biology 4361 Early Development in Invertebrates October 25, 2006 Early Development Overview Cleavage rapid cell divisions divisions of fertilized egg into many cells Gastrulation
Developmental Biology Biology 4361 Early Development in Invertebrates October 25, 2006 Early Development Overview Cleavage rapid cell divisions divisions of fertilized egg into many cells Gastrulation
Designation of the Anterior/Posterior Axis in Pregastrula Xenopus laevis
 Developmental Biology 225, 37 58 (2000) doi:10.1006/dbio.2000.9803, available online at http://www.idealibrary.com on Designation of the Anterior/Posterior Axis in Pregastrula Xenopus laevis Mary Constance
Developmental Biology 225, 37 58 (2000) doi:10.1006/dbio.2000.9803, available online at http://www.idealibrary.com on Designation of the Anterior/Posterior Axis in Pregastrula Xenopus laevis Mary Constance
Question Set # 4 Answer Key 7.22 Nov. 2002
 Question Set # 4 Answer Key 7.22 Nov. 2002 1) A variety of reagents and approaches are frequently used by developmental biologists to understand the tissue interactions and molecular signaling pathways
Question Set # 4 Answer Key 7.22 Nov. 2002 1) A variety of reagents and approaches are frequently used by developmental biologists to understand the tissue interactions and molecular signaling pathways
Role of Organizer Chages in Late Frog Embryos
 Ectoderm Germ Layer Frog Fate Map Frog Fate Map Role of Organizer Chages in Late Frog Embryos Organizer forms three distinct regions Notochord formation in chick Beta-catenin localization How does beta-catenin
Ectoderm Germ Layer Frog Fate Map Frog Fate Map Role of Organizer Chages in Late Frog Embryos Organizer forms three distinct regions Notochord formation in chick Beta-catenin localization How does beta-catenin
Drosophila melanogaster- Morphogen Gradient
 NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
NPTEL Biotechnology - Systems Biology Drosophila melanogaster- Morphogen Gradient Dr. M. Vijayalakshmi School of Chemical and Biotechnology SASTRA University Joint Initiative of IITs and IISc Funded by
Tsukushi Modulates Xnr2, FGF and BMP Signaling: Regulation of Xenopus Germ Layer Formation
 Tsukushi Modulates Xnr2, FGF and BMP Signaling: Regulation of Xenopus Germ Layer Formation Samantha A. Morris 1 *, Alexandra D. Almeida 1, Hideaki Tanaka 2, Kunimasa Ohta 2, Shin-ichi Ohnuma 1 * 1 Department
Tsukushi Modulates Xnr2, FGF and BMP Signaling: Regulation of Xenopus Germ Layer Formation Samantha A. Morris 1 *, Alexandra D. Almeida 1, Hideaki Tanaka 2, Kunimasa Ohta 2, Shin-ichi Ohnuma 1 * 1 Department
Localization of the factors producing the periodic activities responsible for synchronous cleavage in Xenopus embryos
 /. Embryol. exp. Morph. 85, 33-46 (1985) 33 Printed in Great Britain The Company of Biologists Limited 1985 Localization of the factors producing the periodic activities responsible for synchronous cleavage
/. Embryol. exp. Morph. 85, 33-46 (1985) 33 Printed in Great Britain The Company of Biologists Limited 1985 Localization of the factors producing the periodic activities responsible for synchronous cleavage
Zygotic Wnt Activity Is Required for Brachyury Expression in the Early Xenopus laevis Embryo
 Developmental Biology 250, 112 127 (2002) doi:10.1006/dbio.2002.0786 Zygotic Wnt Activity Is Required for Brachyury Expression in the Early Xenopus laevis Embryo Alin Vonica 1 and Barry M. Gumbiner 2 Cellular
Developmental Biology 250, 112 127 (2002) doi:10.1006/dbio.2002.0786 Zygotic Wnt Activity Is Required for Brachyury Expression in the Early Xenopus laevis Embryo Alin Vonica 1 and Barry M. Gumbiner 2 Cellular
Exam 1 ID#: October 4, 2007
 Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Biology 4361 Name: KEY Exam 1 ID#: October 4, 2007 Multiple choice (one point each) (1-25) 1. The process of cells forming tissues and organs is called a. morphogenesis. b. differentiation. c. allometry.
Xenopus mothers against decapentaplegic is an embryonic ventralizing agent
 Development 122, 2359-2366 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV3476 2359 Xenopus mothers against decapentaplegic is an embryonic ventralizing agent that acts downstream
Development 122, 2359-2366 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV3476 2359 Xenopus mothers against decapentaplegic is an embryonic ventralizing agent that acts downstream
Developmental Biology Lecture Outlines
 Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Developmental Biology Lecture Outlines Lecture 01: Introduction Course content Developmental Biology Obsolete hypotheses Current theory Lecture 02: Gametogenesis Spermatozoa Spermatozoon function Spermatozoon
Chapter 10 Development and Differentiation
 Part III Organization of Cell Populations Chapter Since ancient times, people have wondered how organisms are formed during the developmental process, and many researchers have worked tirelessly in search
Part III Organization of Cell Populations Chapter Since ancient times, people have wondered how organisms are formed during the developmental process, and many researchers have worked tirelessly in search
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
9/4/2015 INDUCTION CHAPTER 1. Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology. Fig 1.
 INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
INDUCTION CHAPTER 1 Neurons are similar across phyla Thus, many different model systems are used in developmental neurobiology Fig 1.1 1 EVOLUTION OF METAZOAN BRAINS GASTRULATION MAKING THE 3 RD GERM LAYER
University of Bristol - Explore Bristol Research
 Domingos, P. M., Itasaki, N., Jones, C. M., Mercurio, S., Sargent, M. G., Smith, J. C., & Krumlauf, R. (2001). The Wnt/beta-catenin pathway posteriorizes neural tissue in Xenopus by an indirect mechanism
Domingos, P. M., Itasaki, N., Jones, C. M., Mercurio, S., Sargent, M. G., Smith, J. C., & Krumlauf, R. (2001). The Wnt/beta-catenin pathway posteriorizes neural tissue in Xenopus by an indirect mechanism
!!!!!!!! DB3230 Midterm 2 12/13/2013 Name:
 1. (10 pts) Draw or describe the fate map of a late blastula stage sea urchin embryo. Draw or describe the corresponding fate map of the pluteus stage larva. Describe the sequence of gastrulation events
1. (10 pts) Draw or describe the fate map of a late blastula stage sea urchin embryo. Draw or describe the corresponding fate map of the pluteus stage larva. Describe the sequence of gastrulation events
Questions in developmental biology. Differentiation Morphogenesis Growth/apoptosis Reproduction Evolution Environmental integration
 Questions in developmental biology Differentiation Morphogenesis Growth/apoptosis Reproduction Evolution Environmental integration Representative cell types of a vertebrate zygote => embryo => adult differentiation
Questions in developmental biology Differentiation Morphogenesis Growth/apoptosis Reproduction Evolution Environmental integration Representative cell types of a vertebrate zygote => embryo => adult differentiation
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Exam 2 ID#: November 9, 2006
 Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Biology 4361 Name: KEY Exam 2 ID#: November 9, 2006 Multiple choice (one point each) Circle the best answer. 1. Inducers of Xenopus lens and optic vesicle include a. pharyngeal endoderm and anterior neural
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its
 The Wnt/ -catenin signalling pathway and its") Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Supplementary Figure 1: Mechanism of Lbx2 action on the Wnt/ -catenin signalling pathway. (a) The Wnt/ -catenin signalling pathway and its transcriptional activity in wild-type embryo. A gradient of canonical
Patterns and control of cell motility in the Xenopus gastrula
 Development 125, 1931-1942 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV6326 1931 Patterns and control of cell motility in the Xenopus gastrula Stephan Wacker 1, Anja Brodbeck
Development 125, 1931-1942 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV6326 1931 Patterns and control of cell motility in the Xenopus gastrula Stephan Wacker 1, Anja Brodbeck
Drosophila Life Cycle
 Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Drosophila Life Cycle 1 Early Drosophila Cleavage Nuclei migrate to periphery after 10 nuclear divisions. Cellularization occurs when plasma membrane folds in to divide nuclei into cells. Drosophila Superficial
Mesoderm Formation in Eleutherodactylus coqui: Body Patterning in a Frog with a Large Egg
 Developmental Biology 236, 109 123 (2001) doi:10.1006/dbio.2001.0310, available online at http://www.idealibrary.com on Mesoderm Formation in Eleutherodactylus coqui: Body Patterning in a Frog with a Large
Developmental Biology 236, 109 123 (2001) doi:10.1006/dbio.2001.0310, available online at http://www.idealibrary.com on Mesoderm Formation in Eleutherodactylus coqui: Body Patterning in a Frog with a Large
MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning
 MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning Learning Goals for Lecture 29 4.1 Describe the contributions of early developmental events in the embryo to the formation
MCDB 4777/5777 Molecular Neurobiology Lecture 29 Neural Development- In the beginning Learning Goals for Lecture 29 4.1 Describe the contributions of early developmental events in the embryo to the formation
The induction of anterior and posterior neural genes in Xenopus laevis
 Development 109, 765-774 (1990) Printed in Great Britain The Company of Biologists Limited 1990 765 The induction of anterior and posterior neural genes in Xenopus laevis C. R. SHARPE and J. B. GURDON
Development 109, 765-774 (1990) Printed in Great Britain The Company of Biologists Limited 1990 765 The induction of anterior and posterior neural genes in Xenopus laevis C. R. SHARPE and J. B. GURDON
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Developmental Biology Biology 4361 Axis Specification in Drosophila July 9, 2008 Drosophila Development Overview Fertilization Cleavage Gastrulation Drosophila body plan Oocyte formation Genetic control
Biology 218, practise Exam 2, 2011
 Figure 3 The long-range effect of Sqt does not depend on the induction of the endogenous cyc or sqt genes. a, Design and predictions for the experiments shown in b-e. b-e, Single-cell injection of 4 pg
Figure 3 The long-range effect of Sqt does not depend on the induction of the endogenous cyc or sqt genes. a, Design and predictions for the experiments shown in b-e. b-e, Single-cell injection of 4 pg
PRACTICE EXAM. 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos.
 PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
PRACTICE EXAM 20 pts: 1. With the aid of a diagram, indicate how initial dorsal-ventral polarity is created in fruit fly and frog embryos. No Low [] Fly Embryo Embryo Non-neural Genes Neuroectoderm Genes
Differentiation 82 (2011) Contents lists available at ScienceDirect. Differentiation. journal homepage:
 Contents lists available at ScienceDirect. Differentiation. journal homepage:") Differentiation 82 (2011) 99 107 Contents lists available at ScienceDirect Differentiation journal homepage: www.elsevier.com/locate/diff Inhibition of FGF signaling converts dorsal mesoderm to ventral
Differentiation 82 (2011) 99 107 Contents lists available at ScienceDirect Differentiation journal homepage: www.elsevier.com/locate/diff Inhibition of FGF signaling converts dorsal mesoderm to ventral
Caudalization of neural fate by tissue recombination and bfgf
 Development 121, 4349-4358 (1995) Printed in Great Britain The Company of Biologists Limited 1995 DEV9434 4349 Caudalization of neural fate by tissue recombination and bfgf Wm. Gregory Cox and Ali Hemmati-Brivanlou
Development 121, 4349-4358 (1995) Printed in Great Britain The Company of Biologists Limited 1995 DEV9434 4349 Caudalization of neural fate by tissue recombination and bfgf Wm. Gregory Cox and Ali Hemmati-Brivanlou
Development of Drosophila
 Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
Development of Drosophila Hand-out CBT Chapter 2 Wolpert, 5 th edition March 2018 Introduction 6. Introduction Drosophila melanogaster, the fruit fly, is found in all warm countries. In cooler regions,
2. Fertilization activates the egg and bring together the nuclei of sperm and egg
 2. Fertilization activates the egg and bring together the nuclei of sperm and egg Sea urchins (what phylum?) are models for the study of the early development of deuterostomes (like us, right?). Sea urchin
2. Fertilization activates the egg and bring together the nuclei of sperm and egg Sea urchins (what phylum?) are models for the study of the early development of deuterostomes (like us, right?). Sea urchin
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
β-catenin localization during Xenopus embryogenesis: accumulation at tissue and somite boundaries
 Development 120, 3667-3679 (1994) Printed in Great Britain The Company of Biologists Limited 1994 3667 β-catenin localization during Xenopus embryogenesis: accumulation at tissue and somite boundaries
Development 120, 3667-3679 (1994) Printed in Great Britain The Company of Biologists Limited 1994 3667 β-catenin localization during Xenopus embryogenesis: accumulation at tissue and somite boundaries
Mesoderm induction: from caps to chips
 Mesoderm induction: from caps to chips David Kimelman Abstract Vertebrate mesoderm induction is one of the classical problems in developmental biology. Various developmental biology approaches, particularly
Mesoderm induction: from caps to chips David Kimelman Abstract Vertebrate mesoderm induction is one of the classical problems in developmental biology. Various developmental biology approaches, particularly
Neural crest induction by paraxial mesoderm in Xenopus embryos requires FGF signals
 Development 130, 3111-3124 2003 The Company of Biologists Ltd doi:10.1242/dev.00531 3111 Neural crest induction by paraxial mesoderm in Xenopus embryos requires FGF signals Anne-Hélène Monsoro-Burq*, Russell
Development 130, 3111-3124 2003 The Company of Biologists Ltd doi:10.1242/dev.00531 3111 Neural crest induction by paraxial mesoderm in Xenopus embryos requires FGF signals Anne-Hélène Monsoro-Burq*, Russell
AP Biology Gene Regulation and Development Review
 AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
AP Biology Gene Regulation and Development Review 1. What does the regulatory gene code for? 2. Is the repressor by default active/inactive? 3. What changes the repressor activity? 4. What does repressor
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development June 23, 2009 Concepts Cell-Cell Communication Cells develop in the context of their environment, including: - their immediate
Biology 4361 - Developmental Biology Cell-Cell Communication in Development June 23, 2009 Concepts Cell-Cell Communication Cells develop in the context of their environment, including: - their immediate
Unicellular: Cells change function in response to a temporal plan, such as the cell cycle.
 Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
Spatial organization is a key difference between unicellular organisms and metazoans Unicellular: Cells change function in response to a temporal plan, such as the cell cycle. Cells differentiate as a
DNA can be extracted from the following sample types using this procedure: Archived
 Sample types Principle Safety Equipment and supplies DNA can be extracted from the following sample types using this procedure: concentrated DNA samples (e.g., blood, saliva, non-contact samples) hair
Sample types Principle Safety Equipment and supplies DNA can be extracted from the following sample types using this procedure: concentrated DNA samples (e.g., blood, saliva, non-contact samples) hair
Zebrafish wnt8 and wnt8b share a common activity but are involved in distinct developmental pathways
 Development 121, 1787-1799 (1995) Printed in Great Britain The Company of Biologists Limited 1995 1787 Zebrafish wnt8 and wnt8b share a common activity but are involved in distinct developmental pathways
Development 121, 1787-1799 (1995) Printed in Great Britain The Company of Biologists Limited 1995 1787 Zebrafish wnt8 and wnt8b share a common activity but are involved in distinct developmental pathways
Data Sheet. Azide Cy5 RNA T7 Transcription Kit
 Cat. No. Size 1. Description PP-501-Cy5 10 reactions à 40 µl For in vitro use only Quality guaranteed for 12 months Store all components at -20 C. Avoid freeze and thaw cycles. DBCO-Sulfo-Cy5 must be stored
Cat. No. Size 1. Description PP-501-Cy5 10 reactions à 40 µl For in vitro use only Quality guaranteed for 12 months Store all components at -20 C. Avoid freeze and thaw cycles. DBCO-Sulfo-Cy5 must be stored
Patterning the early Xenopus embryo
 REVIEW 1205 Development 133, 1205-1217 (2006) doi:10.1242/dev.02304 Patterning the early Xenopus embryo Janet Heasman Developmental biology teachers use the example of the frog embryo to introduce young
REVIEW 1205 Development 133, 1205-1217 (2006) doi:10.1242/dev.02304 Patterning the early Xenopus embryo Janet Heasman Developmental biology teachers use the example of the frog embryo to introduce young
NIH Public Access Author Manuscript Int J Dev Biol. Author manuscript; available in PMC 2012 January 19.
 NIH Public Access Author Manuscript Published in final edited form as: Int J Dev Biol. 2011 ; 55(10-11-12): 917 921. doi:10.1387/ijdb.113288sh. XIer2 is required for convergent extension movements during
NIH Public Access Author Manuscript Published in final edited form as: Int J Dev Biol. 2011 ; 55(10-11-12): 917 921. doi:10.1387/ijdb.113288sh. XIer2 is required for convergent extension movements during
Autonomous mesoderm formation in blastocoelic roof explants from inverted Xenopus embryos
 Int. J. De\'. Riol. 36: 115-122 (1992) 115 Autonomous mesoderm formation in blastocoelic roof explants from inverted Xenopus embryos RENEE TENCER' and MARIANNE GOLDFINGER Laboratory of Developmentaf Biology,
Int. J. De\'. Riol. 36: 115-122 (1992) 115 Autonomous mesoderm formation in blastocoelic roof explants from inverted Xenopus embryos RENEE TENCER' and MARIANNE GOLDFINGER Laboratory of Developmentaf Biology,
Chapter 11. Development: Differentiation and Determination
 KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
KAP Biology Dept Kenyon College Differential gene expression and development Mechanisms of cellular determination Induction Pattern formation Chapter 11. Development: Differentiation and Determination
Phenol-Chloroform reagents. Selection guide. OH ; MW : High quality reagents for use in nucleic acid purification.
 Phenol-Chloroform reagents Extraction with phenol and phenol/chloroform mixtures is a universal method for purification of DNA and RNA. Proteins and restriction enzymes are removed by phenol and chloroform
Phenol-Chloroform reagents Extraction with phenol and phenol/chloroform mixtures is a universal method for purification of DNA and RNA. Proteins and restriction enzymes are removed by phenol and chloroform
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11589 Supplementary Figure 1 Ciona intestinalis and Petromyzon marinus neural crest expression domain comparison. Cartoon shows dorsal views of Ciona mid gastrula (left) and Petromyzon
doi:10.1038/nature11589 Supplementary Figure 1 Ciona intestinalis and Petromyzon marinus neural crest expression domain comparison. Cartoon shows dorsal views of Ciona mid gastrula (left) and Petromyzon
Anteroposterior neural tissue specification by activin-induced mesoderm
 Proc. Natl. Acad. Sci. USA Vol. 94, pp. 8596 8601, August 1997 Developmental Biology Anteroposterior neural tissue specification by activin-induced mesoderm JEREMY B. A. GREEN*, T.LES COOK, J.C.SMITH,
Proc. Natl. Acad. Sci. USA Vol. 94, pp. 8596 8601, August 1997 Developmental Biology Anteroposterior neural tissue specification by activin-induced mesoderm JEREMY B. A. GREEN*, T.LES COOK, J.C.SMITH,
MBios 401/501: Lecture 14.2 Cell Differentiation I. Slide #1. Cell Differentiation
 MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
MBios 401/501: Lecture 14.2 Cell Differentiation I Slide #1 Cell Differentiation Cell Differentiation I -Basic principles of differentiation (p1305-1320) -C-elegans (p1321-1327) Cell Differentiation II
posterior end mark, a novel maternal gene encoding a localized factor in the
 Development 122, 2005-2012 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV5066 2005 posterior end mark, a novel maternal gene encoding a localized factor in the ascidian embryo
Development 122, 2005-2012 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV5066 2005 posterior end mark, a novel maternal gene encoding a localized factor in the ascidian embryo
Developmental Biology 3230 Midterm Exam 1 March 2006
 Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Name Developmental Biology 3230 Midterm Exam 1 March 2006 1. (20pts) Regeneration occurs to some degree to most metazoans. When you remove the head of a hydra a new one regenerates. Graph the inhibitor
Later embryogenesis: regulatory circuitry in morphogenetic fields
 Development 118, 665-690 (1993) Printed in Great Britain The Company of Biologists Limited 1993 Review Article 665 Later embryogenesis: regulatory circuitry in morphogenetic fields Eric H. Davidson Division
Development 118, 665-690 (1993) Printed in Great Britain The Company of Biologists Limited 1993 Review Article 665 Later embryogenesis: regulatory circuitry in morphogenetic fields Eric H. Davidson Division
Cell-Cell Communication in Development
 Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Biology 4361 - Developmental Biology Cell-Cell Communication in Development October 2, 2007 Cell-Cell Communication - Topics Induction and competence Paracrine factors inducer molecules Signal transduction
Expression of zebrafish goosecoid and no tail gene products in wild-type and mutant no tail embryos
 Development 120, 843-852 (1994) Printed in Great Britain The Company of Biologists Limited 1994 843 Expression of zebrafish goosecoid and no tail gene products in wild-type and mutant no tail embryos S.
Development 120, 843-852 (1994) Printed in Great Britain The Company of Biologists Limited 1994 843 Expression of zebrafish goosecoid and no tail gene products in wild-type and mutant no tail embryos S.
Mutual antagonism of SoxB1 and. canonical Wnt signaling in sea urchin embryos
 Mutual antagonism of SoxB1 and canonical Wnt signaling in sea urchin embryos Review by Lynne M. Angerer 1,4, Alan P. Kenny 2, Laurel A. Newman 3 and Robert C. Angerer 1 1 National Institute for Dental
Mutual antagonism of SoxB1 and canonical Wnt signaling in sea urchin embryos Review by Lynne M. Angerer 1,4, Alan P. Kenny 2, Laurel A. Newman 3 and Robert C. Angerer 1 1 National Institute for Dental
Protease Inhibitor Cocktail A (1 tablet / 7 10 ml, Roche Cat# ) Protease inhibitor Cocktail B (0.5ml per 250ml, Calbiochem Cat# )
 Protease inhibitor Cocktail B (0.5ml per 250ml, Calbiochem Cat# )") Protocol for Western Blotting Tissue/Cell Sample Preparation Lysis Buffer 1 (ph8.0) o 50mM Tris-Cl o 150mM NaCl o 1% v/v NP40 o protease inhibitor cocktail A/B Lysis Buffer 2 (RIPA) (ph 8.0) o 50mM Tris-Cl
Protocol for Western Blotting Tissue/Cell Sample Preparation Lysis Buffer 1 (ph8.0) o 50mM Tris-Cl o 150mM NaCl o 1% v/v NP40 o protease inhibitor cocktail A/B Lysis Buffer 2 (RIPA) (ph 8.0) o 50mM Tris-Cl
Life Sciences For NET & SLET Exams Of UGC-CSIR. Section B and C. Volume-08. Contents A. BASIC CONCEPT OF DEVELOPMENT 1
 Section B and C Volume-08 Contents 5. DEVELOPMENTAL BIOLOGY A. BASIC CONCEPT OF DEVELOPMENT 1 B. GAMETOGENESIS, FERTILIZATION AND EARLY DEVELOPMENT 23 C. MORPHOGENESIS AND ORGANOGENESIS IN ANIMALS 91 0
Section B and C Volume-08 Contents 5. DEVELOPMENTAL BIOLOGY A. BASIC CONCEPT OF DEVELOPMENT 1 B. GAMETOGENESIS, FERTILIZATION AND EARLY DEVELOPMENT 23 C. MORPHOGENESIS AND ORGANOGENESIS IN ANIMALS 91 0
Developmental processes Differential gene expression Introduction to determination The model organisms used to study developmental processes
 Date Title Topic(s) Learning Outcomes: Sept 28 Oct 3 1. What is developmental biology and why should we care? 2. What is so special about stem cells and gametes? Developmental processes Differential gene
Date Title Topic(s) Learning Outcomes: Sept 28 Oct 3 1. What is developmental biology and why should we care? 2. What is so special about stem cells and gametes? Developmental processes Differential gene
Evolution of the Organizer and the chordate body plan
 Int. J. Dev. Biol. 45: 133-153 (2001) Evolution of the organizer 133 Evolution of the Organizer and the chordate body plan JOHN GERHART* Department of Molecular and Cell Biology. University of California,
Int. J. Dev. Biol. 45: 133-153 (2001) Evolution of the organizer 133 Evolution of the Organizer and the chordate body plan JOHN GERHART* Department of Molecular and Cell Biology. University of California,
CHAPTER 1 INTRODUCTION AXIS FORMATION IN XENOPUS LAEVIS
 CHAPTER 1 INTRODUCTION AXIS FORMATION IN XENOPUS LAEVIS CHAPTER 1 9 AXIS FORMATION During vertebrate embryonic development, a cellular pattern is established, which specifies the formation of the primary
CHAPTER 1 INTRODUCTION AXIS FORMATION IN XENOPUS LAEVIS CHAPTER 1 9 AXIS FORMATION During vertebrate embryonic development, a cellular pattern is established, which specifies the formation of the primary
Axis determination in flies. Sem 9.3.B.5 Animal Science
 Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Axis determination in flies Sem 9.3.B.5 Animal Science All embryos are in lateral view (anterior to the left). Endoderm, midgut; mesoderm; central nervous system; foregut, hindgut and pole cells in yellow.
Endoderm Specification and Differentiation in Xenopus Embryos
 Developmental Biology 236, 330 343 (2001) doi:10.1006/dbio.2001.0347, available online at http://www.idealibrary.com on Endoderm Specification and Differentiation in Xenopus Embryos Marko E. Horb 1 and
Developmental Biology 236, 330 343 (2001) doi:10.1006/dbio.2001.0347, available online at http://www.idealibrary.com on Endoderm Specification and Differentiation in Xenopus Embryos Marko E. Horb 1 and
The role of the zebrafish nodal-related genes squint and cyclops in patterning of mesendoderm
 Development 130, 1837-1851 2003 The Company of Biologists Ltd doi:10.1242/dev.00400 1837 The role of the zebrafish nodal-related genes squint and cyclops in patterning of mesendoderm Scott T. Dougan 1,
Development 130, 1837-1851 2003 The Company of Biologists Ltd doi:10.1242/dev.00400 1837 The role of the zebrafish nodal-related genes squint and cyclops in patterning of mesendoderm Scott T. Dougan 1,
Bio 127 Section I Introduction to Developmental Biology. Cell Cell Communication in Development. Developmental Activities Coordinated in this Way
 Bio 127 Section I Introduction to Developmental Biology Cell Cell Communication in Development Gilbert 9e Chapter 3 It has to be EXTREMELY well coordinated for the single celled fertilized ovum to develop
Bio 127 Section I Introduction to Developmental Biology Cell Cell Communication in Development Gilbert 9e Chapter 3 It has to be EXTREMELY well coordinated for the single celled fertilized ovum to develop
Organizer and axes formation as a self-organizing process
 Int. J. Dev. Biol. 45: 177-188 (2001) Organizer and axes formation as a self-organizing process 177 Organizer and axes formation as a self-organizing process HANS MEINHARDT Max-Planck-Institut für Entwicklungsbiologie,
Int. J. Dev. Biol. 45: 177-188 (2001) Organizer and axes formation as a self-organizing process 177 Organizer and axes formation as a self-organizing process HANS MEINHARDT Max-Planck-Institut für Entwicklungsbiologie,
Endogenous patterns of TGFβ superfamily signaling during early Xenopus development
 Development 127, 2917-2931 (2000) Printed in Great ritain The Company of iologists Limited 2000 DEV4318 2917 Endogenous patterns of TGFβ superfamily signaling during early Xenopus development Sandrine
Development 127, 2917-2931 (2000) Printed in Great ritain The Company of iologists Limited 2000 DEV4318 2917 Endogenous patterns of TGFβ superfamily signaling during early Xenopus development Sandrine
Cell Cell Communication in Development
 Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Biology 4361 Developmental Biology Cell Cell Communication in Development June 25, 2008 Cell Cell Communication Concepts Cells in developing organisms develop in the context of their environment, including
Neural development its all connected
 Neural development its all connected How do you build a complex nervous system? How do you build a complex nervous system? 1. Learn how tissue is instructed to become nervous system. Neural induction 2.
Neural development its all connected How do you build a complex nervous system? How do you build a complex nervous system? 1. Learn how tissue is instructed to become nervous system. Neural induction 2.
Bmp-4 acts as a morphogen in dorsoventral mesoderm patterning in Xenopus
 Development 124, 23-2334 (1997) Printed in Great Britain The Company of Biologists Limited 1997 DEV2158 23 Bmp-4 acts as a morphogen in dorsoventral mesoderm patterning in Xenopus Roland Dosch 1, Volker
Development 124, 23-2334 (1997) Printed in Great Britain The Company of Biologists Limited 1997 DEV2158 23 Bmp-4 acts as a morphogen in dorsoventral mesoderm patterning in Xenopus Roland Dosch 1, Volker
Involvement of the small GTPases XRhoA and XRnd1 in cell adhesion and head formation in early Xenopus development
 Development 126, 5339-5351 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV5327 5339 Involvement of the small GTPases XRhoA and XRnd1 in cell adhesion and head formation in early
Development 126, 5339-5351 (1999) Printed in Great Britain The Company of Biologists Limited 1999 DEV5327 5339 Involvement of the small GTPases XRhoA and XRnd1 in cell adhesion and head formation in early
Spatio-Temporal Expression Patterns of Eight Epidermis-Specific Genes in the Ascidian Embryo
 Spatio-Temporal Expression Patterns of Eight Epidermis-Specific Genes in the Ascidian Embryo Author(s): Kouichi Ishida, Tatsuya Ueki, and Noriyuki Satoh Source: Zoological Science, 13(5):699-709. Published
Spatio-Temporal Expression Patterns of Eight Epidermis-Specific Genes in the Ascidian Embryo Author(s): Kouichi Ishida, Tatsuya Ueki, and Noriyuki Satoh Source: Zoological Science, 13(5):699-709. Published
The Xvent-2 homeobox gene is part of the BMP-4 signalling pathway controling dorsoventral patterning of Xenopus mesoderm
 Development 122, 3045-3053 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV2087 3045 The Xvent-2 homeobox gene is part of the BMP-4 signalling pathway controling dorsoventral
Development 122, 3045-3053 (1996) Printed in Great Britain The Company of Biologists Limited 1996 DEV2087 3045 The Xvent-2 homeobox gene is part of the BMP-4 signalling pathway controling dorsoventral
On the a clock' mechanism determining the time of tissue-specific enzyme development during ascidian embryogenesis
 /. Embryo!, exp. Morph. Vol. 54, pp. 131-139, 1979 Printed in Great Britain Company of Biologists Limited 1979 On the a clock' mechanism determining the time of tissue-specific enzyme development during
/. Embryo!, exp. Morph. Vol. 54, pp. 131-139, 1979 Printed in Great Britain Company of Biologists Limited 1979 On the a clock' mechanism determining the time of tissue-specific enzyme development during
Sonic hedgehog (Shh) signalling in the rabbit embryo
 signalling in the rabbit embryo") Sonic hedgehog (Shh) signalling in the rabbit embryo In the first part of this thesis work the physical properties of cilia-driven leftward flow were characterised in the rabbit embryo. Since its discovery
Sonic hedgehog (Shh) signalling in the rabbit embryo In the first part of this thesis work the physical properties of cilia-driven leftward flow were characterised in the rabbit embryo. Since its discovery
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent
 TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
TrioMol Isolation Reagent Technical Manual No. 0242 Version 06142007 I Description... 1 II Key Features... 1 III Storage..... 1 IV General Protocol Using Triomol Isolation Reagent 1 V Troubleshooting.
Principles of Experimental Embryology
 Biology 4361 Developmental Biology Principles of Experimental Embryology June 16, 2008 Overview What forces affect embryonic development? The embryonic environment: external and internal How do forces
Biology 4361 Developmental Biology Principles of Experimental Embryology June 16, 2008 Overview What forces affect embryonic development? The embryonic environment: external and internal How do forces
7.06 Problem Set
 7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
7.06 Problem Set 5 -- 2006 1. In the first half of the course, we encountered many examples of proteins that entered the nucleus in response to the activation of a cell-signaling pathway. One example of
Developmental Biology Biology 4361
 Developmental Biology Biology 4361 The Anatomical Tradition 2009 A hen is only an egg s way of making a new egg. Samuel Butler, 1885 The Anatomical Tradition - Overview What is developmental biology? How
Developmental Biology Biology 4361 The Anatomical Tradition 2009 A hen is only an egg s way of making a new egg. Samuel Butler, 1885 The Anatomical Tradition - Overview What is developmental biology? How
Nuclearization of β-catenin in ectodermal precursors confers organizer-like ability to induce endomesoderm and pattern a pluteus larva
 Byrum and Wikramanayake EvoDevo 2013, 4:31 RESEARCH Open Access Nuclearization of β-catenin in ectodermal precursors confers organizer-like ability to induce endomesoderm and pattern a pluteus larva Christine
Byrum and Wikramanayake EvoDevo 2013, 4:31 RESEARCH Open Access Nuclearization of β-catenin in ectodermal precursors confers organizer-like ability to induce endomesoderm and pattern a pluteus larva Christine
Regionality of egg cytoplasm that promotes muscle differentiation in embryo of the ascidian, Halocynthia roretzi
 Development 116, 521-529 (1992) Printed in Great Britain The Company of Biologists Limited 1992 521 Regionality of egg cytoplasm that promotes muscle differentiation in embryo of the ascidian, Halocynthia
Development 116, 521-529 (1992) Printed in Great Britain The Company of Biologists Limited 1992 521 Regionality of egg cytoplasm that promotes muscle differentiation in embryo of the ascidian, Halocynthia
Bis sulfone Reagents. Figure 1.
 Bis sulfone Reagents An intact IgG molecule has four accessible inter chain disulfide bonds that can be reduced to form eight free cysteine thiols, which can serve as sites for conjugation. The reaction
Bis sulfone Reagents An intact IgG molecule has four accessible inter chain disulfide bonds that can be reduced to form eight free cysteine thiols, which can serve as sites for conjugation. The reaction
OESTREICH LAB CHIP PROTOCOL
 OESTREICH LAB CHIP PROTOCOL Buffers (all starred (*) buffers must be autoclaved before use) 10X Cell Lysis Buffer*: 100mL 0.214g Potassium Acetate (KOAc) = 10mM 0.147g Magnesium Acetate (MgAc) = 15mM 10mL
OESTREICH LAB CHIP PROTOCOL Buffers (all starred (*) buffers must be autoclaved before use) 10X Cell Lysis Buffer*: 100mL 0.214g Potassium Acetate (KOAc) = 10mM 0.147g Magnesium Acetate (MgAc) = 15mM 10mL
Principles of Experimental Embryology
 Biology 4361 Developmental Biology Principles of Experimental Embryology September 19, 2006 Major Research Questions How do forces outside the embryo affect its development? (Environmental Developmental
Biology 4361 Developmental Biology Principles of Experimental Embryology September 19, 2006 Major Research Questions How do forces outside the embryo affect its development? (Environmental Developmental
Three different fusions led to three basic ideas: 1) If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes
 If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes") Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody
 Based on Monoclonal Anti-cGMP Antibody") (FOR RESEARCH USE ONLY. DO NOT USE IT IN CLINICAL DIAGNOSIS!) cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody Catalog No: E-EL-DS02 96T This manual must be read attentively and
(FOR RESEARCH USE ONLY. DO NOT USE IT IN CLINICAL DIAGNOSIS!) cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody Catalog No: E-EL-DS02 96T This manual must be read attentively and
Kingdom Animalia. Zoology the study of animals
 Kingdom Animalia Zoology the study of animals Summary Animals are multicellular and eukaryotic. consume and digest organic materials thereby being heterotrophs. Most are motile at some time in their lives.
Kingdom Animalia Zoology the study of animals Summary Animals are multicellular and eukaryotic. consume and digest organic materials thereby being heterotrophs. Most are motile at some time in their lives.
RNA Synthesis and Processing
 RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
RNA Synthesis and Processing Introduction Regulation of gene expression allows cells to adapt to environmental changes and is responsible for the distinct activities of the differentiated cell types that
Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8
 Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8 1. Inductive signaling is a hallmark of vertebrate and mammalian development. In early neural development, there are multiple signaling pathways
Cellular Neurobiology BIPN 140 Fall 2016 Problem Set #8 1. Inductive signaling is a hallmark of vertebrate and mammalian development. In early neural development, there are multiple signaling pathways
Behavior of DNA-lacking mitochondria in Entamoeba histolytica revealed by organelle transplant
 9 10 11 Behavior of DNA-lacking mitochondria in Entamoeba histolytica revealed by organelle transplant Makoto Kazama 1 *, Sanae Ogiwara, Takashi Makiuchi 1, Kazuhiro Yoshida 1, Kumiko Nakada-Tsukui, Tomoyoshi
9 10 11 Behavior of DNA-lacking mitochondria in Entamoeba histolytica revealed by organelle transplant Makoto Kazama 1 *, Sanae Ogiwara, Takashi Makiuchi 1, Kazuhiro Yoshida 1, Kumiko Nakada-Tsukui, Tomoyoshi