Phosphatase 2A negatively regulates mitotic exit in
|
|
|
- Morgan Watkins
- 6 years ago
- Views:
Transcription
1 Phosphatase 2A negatively regulates mitotic exit in Saccharomyces cerevisiae Yanchang Wang* and Tuen-Yung Ng Department of Biomedical Sciences, College of Medicine, Florida State University 1115 West Call Street, FL *To whom correspondence should be addressed Fax: Phone: Key words: mitotic exit, phosphatase 2A, Cdc55, yeast. Running title: PP2A inhibits mitotic exit - 1 -
2 Abstract: In budding yeast Saccharomyces cerevisiae, Cdc5 kinase is a component of mitotic exit network (MEN), which inactivates cyclin dependent kinase (CDK) after chromosome segregation. cdc5-1 mutants arrest at telophase at the non-permissive temperature due to the failure of CDK inactivation. To identify more negative regulators of MEN, we carried out a genetic screen for genes that are toxic to cdc5-1 mutants when overexpressed. Genes that encode the B-regulatory subunit (Cdc55) and the three catalytic subunits (Pph21, Pph22 and Pph3) of phosphatase 2A (PP2A) were isolated. In addition to cdc5-1, overexpression of CDC55, PPH21 or PPH22 is also toxic to other temperature sensitive mutants that display defects in mitotic exit. Consistently, deletion of CDC55 partially suppresses the temperature sensitivity of these mutants. Moreover, in the presence of spindle damage, PP2A mutants display nuclear localized Cdc14, the key player in MEN pathway, indicative of MEN activation. All the evidence suggests the negative role of PP2A in mitotic exit. Finally, our genetic and biochemical data suggest that PP2A regulates the phosphorylation of Tem1, which acts at the very top of MEN pathway
3 Introduction: The key driving force for cell division in all eukaryotic cells is the conserved cyclin dependent kinase (CDK). CDK activity fluctuates during the cell cycle, peaking at the S and M phases and dropping at the G1 phase. While the high CDK activity is required for DNA duplication and chromosome segregation, the low CDK activity at late M and G1 phase is essential for the loading of DNA replication complexes onto replication origins (Noton and Diffley, 2000; Lengronne and Schwob, 2002). Thus, cells have developed a signal transduction pathway, named the mitotic exit network (MEN), to inactivate CDK after mitosis (Morgan, 1999). The components of the MEN pathway include protein kinases Cdc5, Cdc15, Dbf2, a GTPase Tem1, a phosphatase Cdc14, and a Dbf2 binding protein Mob1 (Jaspersen et al., 1998; Luca and Winey, 1998; Lee et al., 2001a). Phosphatase Cdc14, the key player in the MEN pathway, dephosphorylates Cdh1, an activator for APC (Anaphase Promoting Complex), and subsequently activates the degradation of a B-type cyclin, Clb2. Cdc14 also enhances the stability of Sic1 protein, which acts as a CDK inhibitor (Visintin et al., 1998). During most of the cell cycle, Cdc14 is localized in the nucleolus, but its translocation out of the nucleolus in anaphase and telophase allows it to encounter its substrates, such as Cdh1 and Sic1. All the components in the MEN pathway are required for the release of Cdc14 from the nucleolus, suggesting that Cdc14 acts downstream of MEN pathway (Shou et al., 1999; Visintin et al., 1999). The regulation of MEN activity is achieved through the multi-layer control of Tem1, a small GTPase that localizes at the spindle pole body (SPB) and acts on the very top of the MEN pathway. Tem1 s activator, the GTPase exchange factor (GEF) Lte1, exhibits daughter-cell - 3 -
4 specific localization. Thus, SPB-localized Tem1 is activated after it encounters Lte1 upon the entrance of SPB into daughter cells (Bardin et al., 2000). Before that, Tem1 is kept inactive by a two-component GTPase activating factor (GAP) composed of Bfa1 and Bub2 (Alexandru et al., 1999; Geymonat et al., 2002; Geymonat et al., 2003). Protein kinase Cdc5 has also been implicated in MEN signaling by regulating Bfa1/Bub2. It phosphorylates one of the GAP components, Bfa1, and frees Tem1 from the inhibition by Bfa1 (Hu et al., 2001; Lee et al., 2001b). Tem1-GTP is believed to activate a downstream protein kinase Cdc15, which then activates the protein kinase Dbf2 in a manner dependent on the Dbf2 associated factor Mob1 (Komarnitsky et al., 1998; Mah et al., 2001). We have identified a new mechanism that inactivates MEN through the induction of Amn1 protein upon MEN activation. Amn1 binds to Tem1 and abolishes its association with the downstream target Cdc15 (Wang et al., 2003). Thus the cooperation of Tem1 s cellular localization, Bfa1/Bub2 GAP activity, and the cell cycle regulated appearance of Amn1, limits the functional window of MEN to late M and early G1 phase. Protein phosphatase 2A regulates a significant array of cellular events. This holoenzyme consists of a catalytic subunit, C, and two regulatory subunits, A and B (Millward et al., 1999). In budding yeast, PPH21, PPH22, and PPH3 encode the catalytic subunits of PP2A, and CDC55 encodes one of the regulatory B subunits (Healy et al., 1991; Ronne et al., 1991; Evans and Stark, 1997). In the presence of nocodazole, a drug that disrupts spindle structures, sister chromatids separate in Δcdc55 mutants, suggesting a function for PP2A in anaphase entry (Minshull et al., 1996; Wang and Burke, 1997). The phenotype of Δcdc55 mutants seems to be the result of increased Cdc28 phosphorylation. In S. cerevisiae, the CDK kinase activity is inhibited in G1 and early S phase by Swe1 kinase dependent phosphorylation at tyrosine 19 of - 4 -
5 Cdc28 (Booher et al., 1993). Δcdc55 mutants are unable to keep a high CDK activity in nocodazole treated cells due to the inhibitory phosphorylation of Cdc28. The premature sister chromatid separation and the cold sensitivity phenotypes in Δcdc55 mutants are suppressed by the CDC28F19 mutation, in which Cdc28 is resistant to the phosphorylation by Swe1 (Minshull et al., 1996; Wang and Burke, 1997). Consistently, Swe1 protein level is increased in Δcdc55 mutants due to the compromised Swe1 protein degradation (Yang et al., 2000). Thus, the accumulation of Swe1 in Δcdc55 mutants results in Cdc28 phosphorylation, which may contribute to the known phenotypes of Δcdc55 mutants. Here we report that PP2A is also involved in mitotic exit regulation. Overproduction of PP2A components is toxic to temperature sensitive mutants that have defects in mitotic exit. Δcdc55 mutants exit mitosis in the presence of spindle damage as judged by the appearance of extra buds and nuclear localized Cdc14. PP2A and Bfa1/Bub2 may negatively regulate mitotic exit in parallel pathways as Δcdc55 Δbfa1 and Δcdc55 Δbub2 double mutants are synthetic sick and exhibit more frequent nuclear localized Cdc14. PP2A may regulate mitotic exit by promoting Tem1 protein dephosphorylation. Thus, we identified a new layer of regulation for mitotic exit, involving PP2A. Results: Genetic screen for genes lethal to cdc5-1 when overexpressed Cdc5 kinase promotes mitotic exit by phosphorylating Bfa1 and leading to its disassociation from Tem1 (Hu et al., 2001). To further understand the regulation of mitotic exit, we carried out a genetic screen for genes that cause lethality in cdc5-1 Ts mutants when overexpressed (Wang et al., 2003). A P GAL -cdna library was introduced into cdc5-1 mutants - 5 -
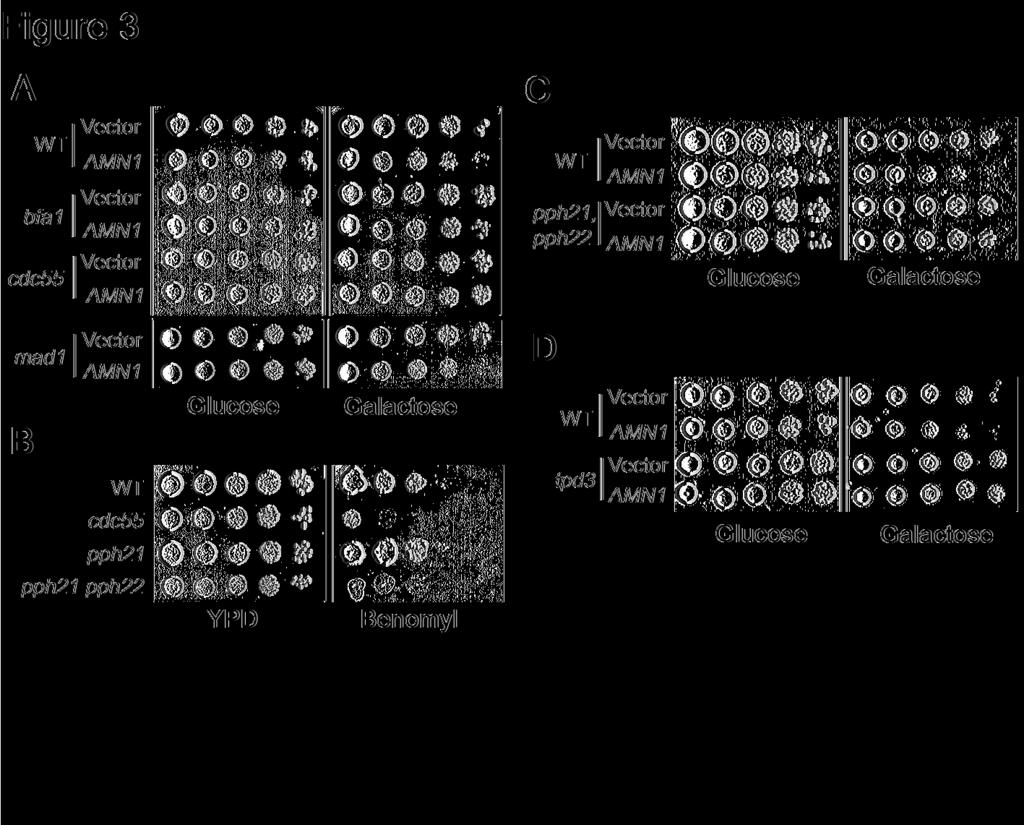
6 and the transformants were selected on URA dropout plates (Liu et al., 1992). From total 12,000 transformants, 10 plasmids exhibited a high dosage lethal phenotype in cdc5-1 mutants. The recovered plasmids contain AMN1, CLB1, CDC55, PPH21, PPH22, and PPH3 genes. The negative role of AMN1 in mitotic exit regulation was reported previously (Wang et al., 2003). Like Clb2, Clb1 could be another APC Cdh1 substrate and its overproduction will deteriorate the growth of cdc5-1 mutants in which the function of APC Cdh1 is compromised (Charles et al., 1998; Shirayama et al., 1998). In addition to AMN1 and CLB1, we also isolated several genes that encode the subunits of PP2A, including the B-regulatory subunit Cdc55 and the catalytic subunits Pph21, Pph22 and Pph3. Cdc5 kinase phosphorylates Bfa1 and promotes mitotic exit (Hu et al., 2001). Also it phosphorylates cohesin Scc1 and facilitates its cleavage by the separase Esp1 (Alexandru et al., 2001). The lethality in cdc5-1 mutants caused by the overexpression of PP2A subunits may result from the negative effects of PP2A on mitotic exit or on other Cdc5 related cell cycle processes. Thus we examined if overexpression of PP2A components is toxic to other temperature sensitive mutants of the MEN pathway. P GAL -CDC55, P GAL -PPH21, P GAL -PPH22, P GAL -PPH3 and a control vector were introduced into tem1-3 and mob1-77 mutants that have defects in mitotic exit (Shirayama et al., 1994; Luca and Winey, 1998). The growth of the transformants was examined after incubation on both glucose and galactose plates at the permissive temperature. Overexpression of CDC55, PPH21 and PPH22 were lethal not only to cdc5-1, but also to tem1-3 and mob1-77 (Figure 1A and data not shown). As overexpression of PP2A components is toxic to the Ts mutants that have defects in mitotic exit, it is likely that PP2A plays a negative role in mitotic exit. In our assays, overexpression of PPH3 was toxic to cdc5-1 mutants, but not to tem1-3 and mob1-77 (data not known). We also noticed that high - 6 -
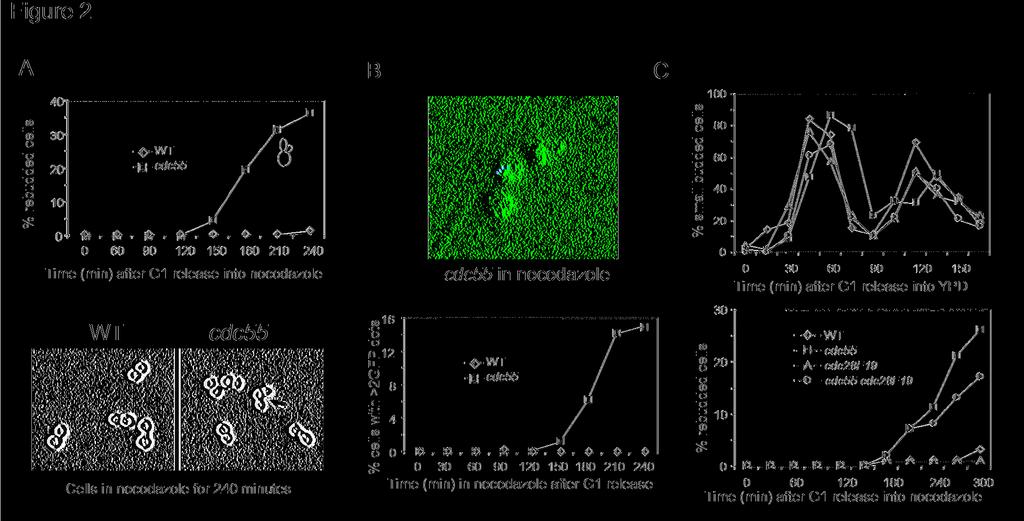
7 dosages of CDC55, PPH21, PPH22, but not PPH3 resulted in slow growth of wild type cells (Figure 1A). If PP2A negatively regulates mitotic exit, inactivation of PP2A might suppress the defects in MEN temperature sensitive mutants. To test this possibility, we generated tem1-3 Δcdc55, cdc15-2 Δcdc55, and cdc5-1 Δcdc55 double mutants and examined their growth at different temperatures. We found that tem1-3 Δcdc55 and cdc5-1 Δcdc55 double mutants were less temperature sensitive than the corresponding single mutants (Figure 1B and 1C). But cdc15-2 and cdc15-2 Δcdc55 mutants exhibited similar growth when incubated at various temperatures. To further confirm the suppression of Ts phenotype of tem1-3 mutants by Δcdc55 deletion, we examined the plating efficiency of tem1-3 and tem1-3 Δcdc55 mutants. The saturated cultures of the two strains were spread onto YPD plates and incubated at 33 C for 14 hours. We found that 98% of the tem1-3 single mutants were arrested as large budded cells, whereas 48% of tem1-3 Δcdc55, mutants formed mini colonies (>4 cells). As high dosages of PP2A regulatory and catalytic subunits are lethal in Ts mutants that are defective in mitotic exit, and deletion of CDC55 partially suppresses the temperature sensitivity of some MEN Ts mutants, we conclude that PP2A plays a negative role in mitotic exit. Δcdc55 mutants exhibit premature mitotic exit phenotype. Bfa1 and Bub2 are required for preventing mitotic exit, and Δbfa1 or Δbub2 mutants rebud in the presence of spindle disruption (Hoyt et al., 1991; Li, 1999). If Cdc55 plays a negative role in mitotic exit, we expect that Δcdc55 mutants will exhibit a similar phenotype. Thus, we examined the cell cycle progression of Δcdc55 mutants in the presence of nocodazole, a microtubule-depolymerizing drug that disrupts the spindle structure. G1 arrested wild-type and - 7 -
8 Δcdc55 mutant cells were released into 30 C YPD medium containing 20μg/ml nocodazole. In wild type cells, disruption of the spindle structure activates the spindle checkpoint and arrests cells at metaphase. After incubation for 3 hours, however, Δcdc55 mutant cells began to rebud. After 4 hour incubation in the presence of nocodazole, about 30% of Δcdc55 mutant cells exhibited extra buds while wild type cells were still arrested as large budded cells, indicating that Δcdc55 mutant cells might exit mitosis (Figure 2A). We also generated Δcdc55 mutants with GFP marked chromosome V to examine sister chromatid separation and chromosome reduplication (Michaelis et al., 1997). As reported previously, a significant portion of Δcdc55 mutant cells showed separated chromatids in the presence of nocodazole. After 4 hour incubation, we noticed that about 10% Δcdc55 mutant cells contained more than two GFP dots, indicating that chromosomes were reduplicated (Figure 2B). The results suggest that Δcdc55 mutant cells are able to exit mitosis and finish the second round of DNA replication in the presence of nocodazole. The premature mitotic exit phenotype in cdc55 mutants is independent of Swe1 Δcdc55 mutant cells exhibit increased Cdc28 phosphorylation at tyrosine 19 (Minshull et al., 1996). Moreover, CDC28F19 mutant that lacks inhibitory phosphorylation site on Cdc28 suppress the cold sensitivity and premature sister separation in Δcdc55 mutants (Wang and Burke, 1997; Yang et al., 2000). Therefore, we tested if the hyperphosphorylation of Cdc28 protein in Δcdc55 mutants also contributes to its premature mitotic exit phenotype. For this purpose, we generated Δcdc55 CDC28F19 double mutants. As reported earlier, the double mutants did not display abnormal bud morphology. To determine if the double mutants exit mitosis, G1 synchronized wild-type, CDC28F19, Δcdc55, and Δcdc55 CDC28F19 cells were - 8 -
9 released into YPD medium containing 20μg/ml nocodazole and the budding indexes were determined. Interestingly, similar to Δcdc55 single mutants, Δcdc55 CDC28F19 double mutants also began rebud after incubation in the presence of nocodazole for 3 hours (Figure 2C bottom). The examination of unperturbed cell cycle progression in these mutants did not show any dramatic discrepancy except that Δcdc55 mutant exhibited slower cell cycle progression (Figure 2C top). We also found that Δswe1 Δcdc55 double mutants were able to rebud in the presence of nocodazole (data not shown). This results argue against the hypothesis that increased phosphorylation of Cdc28 by Swe1 contributes to the premature mitotic exit in Δcdc55 mutants. Δcdc55 mutant suppresses AMN1 overexpression phenotype Overexpression of AMN1 gene slows down cell cycle progression because of its inhibition of mitotic exit (Wang et al., 2003). We predicted that AMN1 overexpression phenotype would be alleviated in mutants with hyperactive MEN. To test this, a vector and a P GAL -AMN1 plasmid were transformed into wild type, Δbfa1, Δbub2, Δmad1, and Δcdc55 mutants. As expected, cells with P GAL -AMN1 plasmid grew slowly on the plates with galactose due to the slower mitotic exit. The AMN1 overexpression phenotype was suppressed by Δbfa1 or Δbub2 deletion, consistent with the negative role of Bfa1/Bub2 complex in mitotic exit regulation (Figure 3A). Similarly, we found that the Δcdc55 mutant also suppressed the AMN1 overexpression phenotype (Figure 3A). However, deletion of MAD1, a spindle checkpoint gene that acts in a different branch from Bfa1/Bub2, could not suppress the AMN1 overexpression phenotype. In response to spindle damage, Mad1, together with other spindle checkpoint components, prevents the activation of APC Cdc20 (Hwang et al., 1998), while Bfa1/Bub2 complex inhibits mitotic exit by keeping Tem1 from activation (Alexandru et al., 1999). Since - 9 -
10 Δbfa1, Δbub2, and Δcdc55 mutants are all able to suppress the AMN1 overexpression phenotype, it is likely that mitotic exit pathways are up-regulated in Δcdc55 mutants. Loss of function of PP2A leads to mitotic exit in cdc55 mutants Cdc55 may function as a negative regulator of PP2A, because the accumulation of Swe1 protein in Δcdc55 mutants is suppressed when PPH21 and PPH22 are deleted (Yang et al., 2000). Therefore, it is not clear whether the loss or the gain of function of PP2A in Δcdc55 mutants leads to the premature mitotic exit. As overexpression of either Cdc55 or catalytic subunits Pph21, Pph22 is toxic to mutants in MEN pathway, it is likely that both Cdc55 and the catalytic subunits of PP2A negatively regulates mitotic exit. If that is the case, mutations in PP2A catalytic subunits will result in a Δcdc55-like phenotype. Previous results indicate that Δcdc55 is sensitive to microtubule disassembly drugs. We therefore examined the growth of Δpph21 single and Δpph21 Δpph22 double mutants on plates containing 15μg/ml benomyl, a microtubule depolymerizing drug like nocodazole. Δpph21 single mutants exhibited similar benomyl sensitivity as wild-type cells, but Δpph21 Δpph22 double mutants failed to form colonies on benomyl plates (Figure 3B). Since both Δcdc55 and Δpph21 Δpph22 mutants are sensitive to microtubule-disassembling drugs, the inactive PP2A in Δcdc55 mutants might contribute to its sensitivity to benomyl. We have shown that Δcdc55 mutants suppress the AMN1 overexpression phenotype, presumably due to the hyper-active mitotic exit pathways. Δpph21 Δpph22 double mutants should alleviate AMN1 overexpression phenotype as well if mitotic exit pathways are hyper active in the double mutants. Therefore, a vector and a P GAL -AMN1 were introduced into Δpph21 Δpph22 double mutants and the growth of the transformants was examined on plates
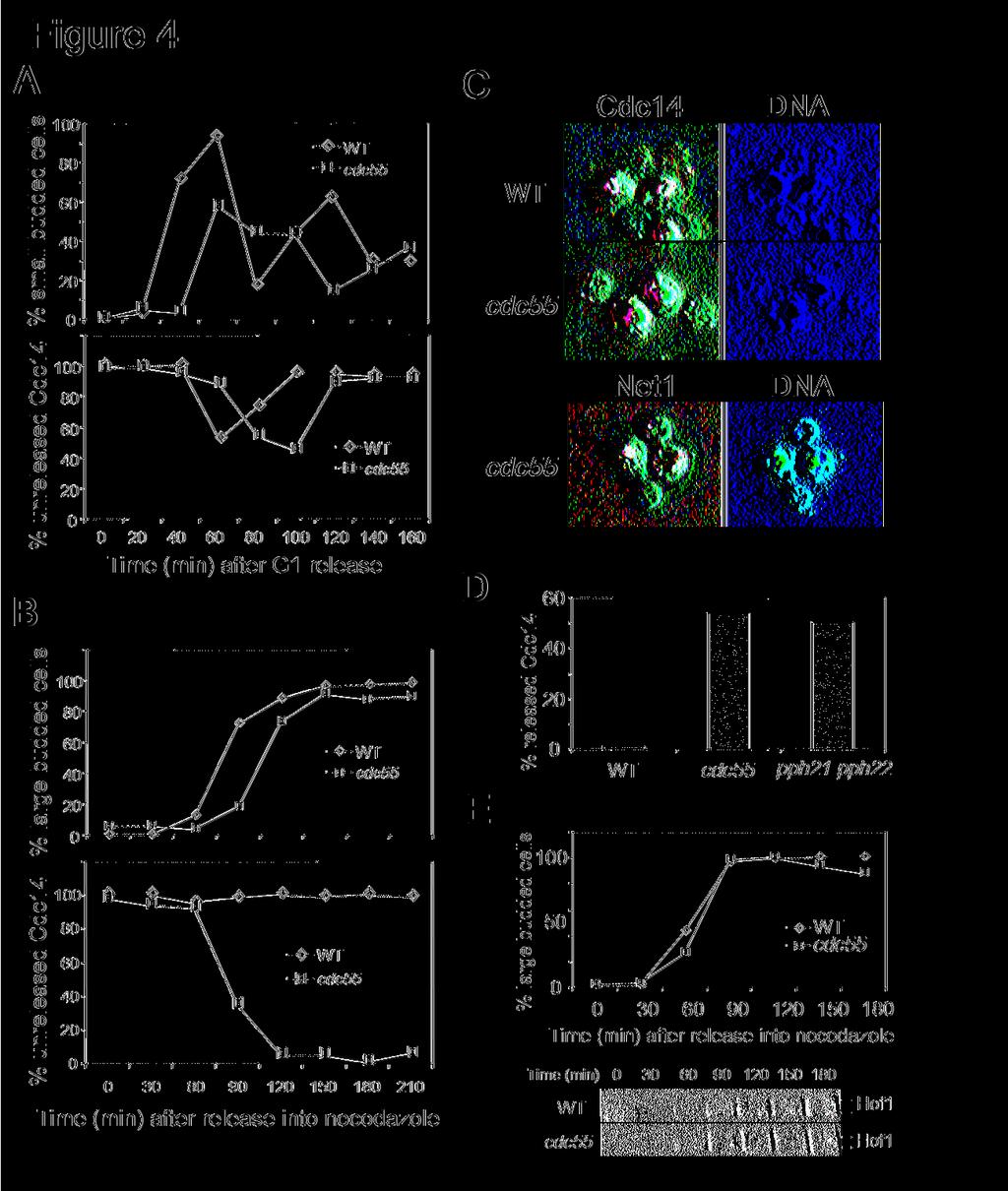
11 containing either glucose or galactose. Δpph21 Δpph22 mutants containing P GAL -AMN1 grew much better than wild-type cells (Figure 3C). Similarly, we examined if loss of the A regulatory subunit (Tpd3) of PP2A also exhibited hyperactive MEN activity and we found that deletion of TPD3 also suppressed AMN1 overexpression phenotype (Figure 3D). Thus, we reason that the regulatory and catalytic subunits of PP2A negatively regulate mitotic exit and the premature mitotic exit phenotype in Δcdc55 mutants results from the loss of function of PP2A. PP2A regulates Cdc14 localization Phosphotase Cdc14 localizes in the nucleolus during most of the cell cycle (Shou et al., 1999; Visintin et al., 1999). After MEN activation, Cdc14 is released into the nucleus so that Cdc14 is able to dephosphorylate its substrates and promote the inactivation of CDK (Visintin et al., 1998). Thus, the localization of Cdc14 has been used as a molecular marker for mitotic exit. We tested the possibility that PP2A inhibits mitotic exit through the regulation of Cdc14 localization. The localization of Cdc14 in wild-type and Δcdc55 mutant cells was examined in the presence of nocodazole. CDC14-HA and Δcdc55 CDC14-HA strains in mid-log phase were synchronized at G1 and then released into 30 C YPD medium containing 20μg/ml nocodazole. The cells were harvested and subjected to immnofluorescence staining. As expected, the majority of wild type cells exhibited nucleolar localized Cdc14, indicating that mitotic exit pathways were inactive. In contrast, Δcdc55 mutant cells showed nuclear localized Cdc14 beginning at 90 minutes after G1 release. After 120 minutes, almost all of the Δcdc55 mutant cells showed nuclear localized Cdc14 in the presence of nocodazole (Figure 4B and 4C). However, in the absence of nocodazole, Δcdc55 mutant exhibited normal cell cycle regulated Cdc14 localization, except that Δcdc55 mutants showed slower cell cycle progression (Figure
12 4A). The nuclear localized Cdc14 in Δcdc55 mutants in the presence of nocodazole could be a result of deformed nucleolar structure. To clear this issue, we examined the localization of Net1, a protein localized in the nucleolus through the cell cycle. G1 synchronized NET1-myc and Δcdc55 NET1-myc cells were released into YPD medium containing nocodazole. In contrast to Cdc14, both wild type and Δcdc55 mutant cells exhibited nucleolar localized Net1 protein in the presence of nocodazole (Figure 4C bottom), suggesting that the nuclear localized Cdc14 in Δcdc55 mutants is not a result of deformed nucleolus. Thus, Δcdc55 mutants fail to keep mitotic exit pathways inactive in the presence of nocodazole and the premature mitotic exit in Δcdc55 mutants is resulted from Cdc14 release from the nucleolus. We also examined the localization of Cdc14 in Δpph21 Δpph22 double mutants in the presence of nocodazole. Asynchronized cells were incubated in YPD medium containing 20μg/ml of nocodazole for 3 hours and Cdc14 localization was examined. Like Δcdc55 mutants, 50% of pph21 pph22 double mutant cells exhibited nuclear localized Cdc14, while almost all the wild type cells showed nucleolar localized Cdc14 (Figure 4D). These data support our conclusion that the B regulatory subunit Cdc55 and the catalytic subunits Pph21 and Pph22 act together to prevent the activation of mitotic exit pathways in the presence of nocodazole. To determine the mitotic exit in molecular level, we analyzed the phosphorylation of Hof1 in wild type and Δcdc55 mutants in the presence of nocodazole. Hof1 is a phosphoprotein required for cytokinesis. The phosphorylation of Hof1 depends on the functional MEN pathway as its phosphorylation is blocked in dbf2-2, cdc14-1, and cdc15-2 mutants (Vallen et al., 2000). Thus, the phosphorylation status of Hof1 protein could be used as a marker of MEN activation. HOF1-HA and Δcdc55 HOF-HA strains were arrested at G1 and then released into YPD medium containing nocodazole. Δcdc55 mutants show more phosphorylated Hof1 protein than wild type
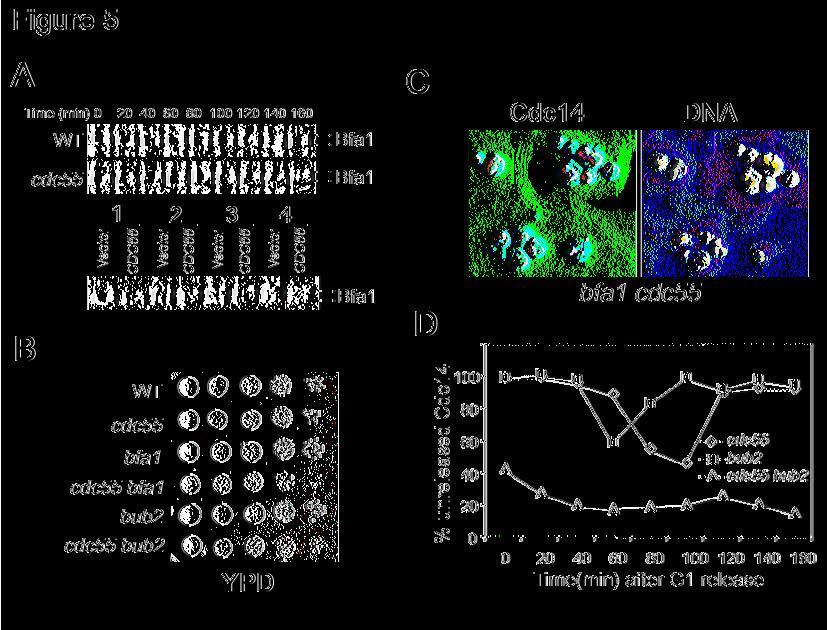
13 cells in the presence of nocodazole, supporting the notion that MEN pathway is hyper-active in Δcdc55 mutants (Figure 4E). PP2A controls mitotic exit independent of Bfa1/Bub2 Our data indicate that PP2A plays a negative role in mitotic exit and Bfa1/Bub2 does so as well. We have demonstrated that protein kinase Cdc5 phosphorylates Bfa1 and promotes mitotic exit (Hu et al., 2001). One reasonable model is that PP2A dephosphorylates Bfa1 and keeps Bfa1 active. If that is the case, mutations in PP2A will result in the hyper-phosphorylation and inactivation of Bfa1. To test this model, we first constructed a Δcdc55 BFA1-HA strain and the phosphorylation of Bfa1 protein was examined in synchronized wild-type and Δcdc55 mutant cells. As with wild type cells, Δcdc55 mutants exhibited cell cycle regulated Bfa1 phosphorylation and increased hyperphosphorylated Bfa1 was not observed in Δcdc55 mutants (Figure 5A). We also examined the phosphorylation of Bfa1 in cells overexpressing CDC55. The results indicate that CDC55 overexpression does not change the Bfa1 phosphorylation profiles (Figure 5A). Therefore, PP2A does not appear to inhibit mitotic exit through dephosphorylation of Bfa1. If Bfa1 is not a substrate of PP2A, it is possible that PP2A regulates mitotic exit in a pathway independent of Bfa1/Bub2. If so, we would expect some synthetic phenotypes when both pathways are abolished. Thus, Δbfa1 Δcdc55 and Δbub2 Δcdc55 double mutants were constructed. Compared to the single mutants, the double mutants exhibited a poor growth phenotype (Figure 5B). Furthermore, in asynchronized cell cultures, about 20% of the double mutants showed extra buds (rebudding) phenotype, indicating that the double mutant cells may exit mitosis prematurely. We then examined the localization of Cdc14 in Δbfa1 Δcdc55 double
14 mutants and found that a significant portion of the double mutant cells exhibited nuclear localized Cdc14 and many double mutant cells had more than one nucleus (Figure 5C). Δcdc55 mutant exhibits SWE1 dependent abnormal morphology. The synthetic phenotype between Δcdc55 and Δbfa1, Δbub2 may come from the combination of abnormal morphology and the spindle checkpoint defects. To test this, we constructed Δcdc55 Δbub2 Δswe1 triple mutants. Even though Δswe1 suppressed the abnormal bud morphology, the synthetic slow growth phenotype of Δcdc55 Δbub2 could not be suppressed by the absence of SWE1. This observation indicates that SWE1 dependent abnormal morphology in Δcdc55 mutant does not contribute to the poor growth phenotype of Δcdc55 Δbfa1 and Δcdc55 Δbub2 double mutants, consistent with the notion that Swe1 accumulation in Δcdc55 mutants is not related to mitotic exit regulation. We also examined the cell cycle regulated localization of Cdc14 in synchronized Δcdc55 Δbub2 Δswe1 triple mutants. As the mutant cells are very sick, only some of the cells responded to α-factor treatment. Thus, only the cells with shmoo morphology were counted for this experiment. We failed to observe proper cell cycle regulated Cdc14 localization in the triple mutants. Even in the G1 arrested cells, about 40% of the triple mutant cells exhibited nucleolar localized Cdc14, while almost all the Δcdc55 and Δbub2 single mutant cells exhibited nucleolar localized Cdc14 (Figure 5D). Based on these results, we conclude that the synthetic phenotype between CDC55 and BFA1/BUB2 is likely due to constitutively activated mitotic exit pathways. The results also suggest that PP2A and Bfa1/Bub2 control mitotic exit in an independent manner and the presence of either one of them is sufficient for a successful mitosis. FEAR pathway may not be required for the mitotic exit in Δcdc55 mutants
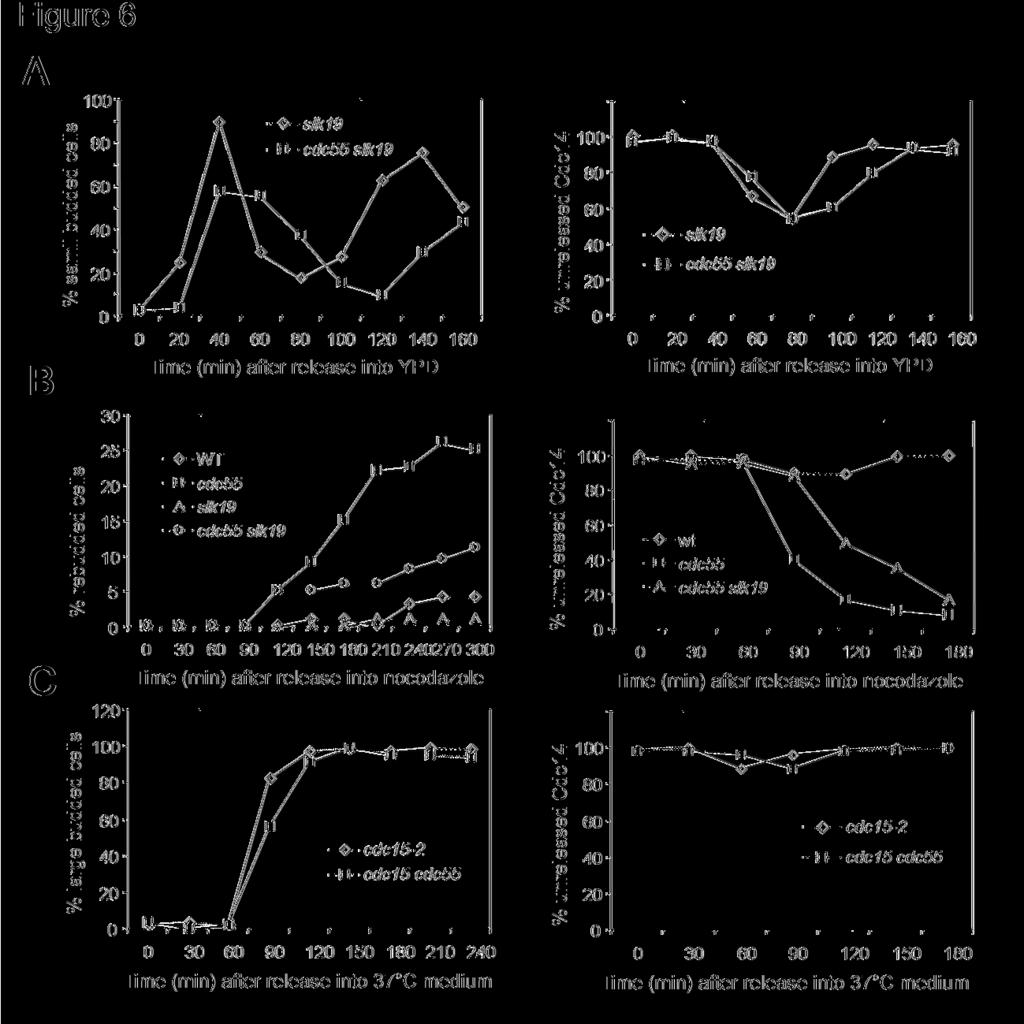
15 What is the target of PP2A that is related to mitotic exit? Both FEAR and MEN pathways control mitotic exit by regulating Cdc14 localization. The FEAR (Cdc fourteen early anaphase release) network promotes Cdc14 release from the nucleolus during early anaphase (Stegmeier et al., 2002). We next addressed the possibility that PP2A regulates mitotic exit by inhibiting the FEAR pathway. If the premature mitotic exit phenotype in Δcdc55 mutants is a result of hyperactive FEAR, deletion of SLK19, which encodes one of the FEAR components, should suppress the Δcdc55 mutant phenotype. We generated Δcdc55 Δslk19 double mutants to examine their rebudding phenotype in the presence of nocodazole. It appeared that the rebudding phenotype of Δcdc55 in the presence of nocodazole was partially suppressed by the Δslk19 mutation (Figure 6B). After 5 hour incubation in the presence of nocodazole, about 20% of Δcdc55 single mutant cells exhibited rebudding morphology. However, only 10% of Δcdc55 Δslk19 double mutant cells rebudded. Since the budding index could not give us a clear-cut answer, we further analyzed mitotic exit in Δcdc55Δ slk19 mutants by examining the localization of Cdc14. G1 arrested wild type, Δcdc55, and Δcdc55 Δslk19 mutants cells with HA tagged CDC14 were released into YPD medium either with or without 20μg/ml of nocodazole. In the absence of nocodazole, Δslk19 and Δslk19 Δcdc55 mutants exhibited cell cycle regulated Cdc14 localization (Figure 6A). In the presence of nocodazole, Cdc14 localized in the nucleus in wild type cells, while Δcdc55 mutants exhibited nuclear localized Cdc14. Δcdc55 Δslk19 double mutants also showed nuclear localized Cdc14, but with delayed kinetics (Figure 6B right). It appeared that defects in FEAR pathway delayed the mitotic exit process in Δcdc55 mutants. One explanation is that PP2A inhibits mitotic exit partially through its inhibition of FEAR pathway. Alternately, the premature release of Cdc14 in Δcdc55 mutants has nothing to do with FEAR pathway, but the defects of this
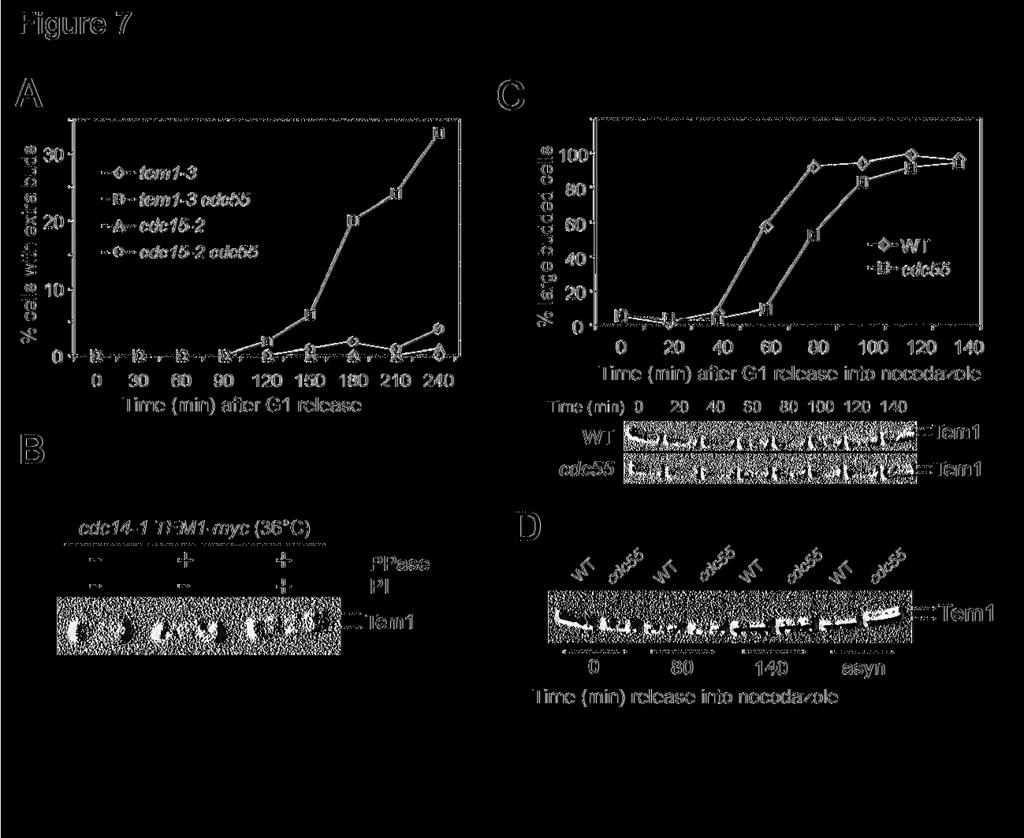
16 pathway slow down the mitotic exit process in Δcdc55 mutants. Thus PP2A must regulate mitotic exit pathway other than FEAR. PP2A and MEN pathway We then tested if MEN was a target of PP2A. If hyperactive MEN pathway contributes to mitotic exit in Δcdc55 mutants, inactivation of MEN components should block the mitotic exit in Δcdc55 mutations. Cdc15 is a component of MEN and cdc15-2 mutants arrest at telophase when incubated at the restrictive temperature (Visintin and Amon, 2001). Thus, we examined the cell cycle progression of cdc15-2 single and cdc15-2 Δcdc55 double mutants to see if the defective MEN could block the mitotic exit in Δcdc55 mutants. G1 arrested cells were released into YPD medium at 37 C and the budding indexes were determined. Both cdc15-2 single and cdc15-2 Δcdc55 double mutants arrested at large budded cells (Figure 6C). Moreover, Cdc14 localized in the nucleolus in both cdc15-2 single and cdc15-2 Δcdc55 double mutants when incubated at the restrictive temperature (Figure 6C), indicating that mitotic exit in Δcdc55 mutants depends upon MEN function. Therefore, we reason that PP2A might negatively regulate the MEN pathway. PP2A may regulate Tem1 protein phosphorylation To answer if the mitotic exit in Δcdc55 mutants depends upon MEN functions, the cell cycle progression of tem1-3 and tem1-3 Δcdc55 was also examined at 37 C. Unlike cdc15-2, the mitotic exit defects in tem1-3 mutants were partially suppressed by deletion of CDC55. After 4 hours incubation, more than 30% of tem1-3 Δcdc55 double mutants exited mitosis as indicated by the appearance of extra buds; however all the cdc15-2 Δcdc55 double mutants were arrested
17 as large budded cells (Figure 7A). This result is consistent with our observation that Δcdc55 mutation partially suppresses the temperature sensitivity of tem1-3 mutants when incubated at 30 C (Figure 1B). As Δcdc55 suppresses the mitotic exit defects in tem1-3 mutants, PP2A might function as a negative regulator of Tem1. It has been shown that Tem1 exhibits cell cycle regulated modification, but the nature of this modification remains unclear. Therefore, we first examined if phosphorylation contributes to the band shift of Tem1 protein. Protein samples were prepared with cdc14-1 TEM1-myc strain incubated at 36 C for two hours, as Tem1 exhibited more slow-migrating forms in cdc14-1 arrested cells. After immunoprecipitation with anti-myc antibody, Tem1 protein was subjected to λ protein phosphatase treatment in the presence or absence of phosphatase inhibitors. We found that the majority of the slow-migrating forms of Tem1 disappeared after λ phosphatase treatment (Figure 7B), indicating that Tem1 is a phosphoprotein. Then we asked if the Tem1 protein phosphorylation is regulated by PP2A. We examined the phosphorylation status of Tem1 in wild type and Δcdc55 mutant cells, and it was very clear that Δcdc55 mutants exhibited more modified Tem1 protein in asynchronized cells (Figure 7D). The phosphorylation of Tem1 was also examined in synchronized TEM1-13myc and Δcdc55 TEM1-13myc cells. G1 arrested cells were released into YPD medium containing 20μg/ml nocodazole at 30 C. In G1 arrested cells, we observed more phosphorylated Tem1 in Δcdc55 mutants. As cells entered S-phase, there were less phosphorylated Tem1 proteins. We noticed the appearance of slow-migrating Tem1 band in Δcdc55 mutants at 100 min; however, the phosphorylated Tem1 did not appear until 140 minutes after G1 release in wild type cells (Figure 7C). When we run the protein samples from wild type and Δcdc55 mutant cells side by side, it was clear that Δcdc55 mutants exhibited more phosphorylated Tem1 protein under various
18 conditions (Figure 7D), indicating that loss of function of PP2A enhances Tem1 phosphorylation. Given the fact that Cdc55 is a component of phosphatase, a reasonable model is that PP2A dephosphorylates Tem1 and inhibits its functions in mitotic exit. Discussion: Previous data indicate that budding yeast Cdc55 regulates bud morphology and sister chromatid separation. Here we report that PP2A also plays a negative role in mitotic exit and several pieces of evidence support this conclusion. First, overexpression of some of the PP2A components is toxic to mutants in which the MEN pathway is compromised. Second, Δcdc55 mutants exhibit extra buds and nuclear localized Cdc14 in the presence of nocodazole. Moreover, the Δcdc55 deletion mutant could suppress the toxicity resulting from the overproduction of Amn1, which acts as a negative regulator of MEN. Finally, Hof1, a protein required for cytokinesis and its phosphorylation depends on activated MEN pathway, exhibits more phosphorylated forms in Δcdc55 mutants in the presence of nocodazole. These results are consistent with our earlier observation that Δcdc55 mutants suppress the temperature sensitivity of cdc20-1 mutants (Wang and Burke, 1997). HCT1/CDH1 was identified as a high copy suppressor of cdc20-1 mutants (Schwab et al., 1997). We reason that the hyper active MEN pathway in Δcdc55 mutants activates APC Cdh1, which overcomes the requirement of Cdc20. It has been shown that Swe1 protein accumulates in Δcdc55 mutants and either Δswe1 deletion or CDC28F19 mutation can suppress the abnormal bud morphology in Δcdc55 mutants (Wang and Burke, 1997; Yang et al., 2000). However, neither Δswe1 deletion nor CDC28F19 mutation could suppress the mitotic exit in Δcdc55 mutants in the presence of nocodazole. Other observations also argue against the role of Swe1 in mitotic exit. We noticed that Δcdc55 Δswe1-18 -
19 double mutant cells are capable of reduplicating their chromosomes in the presence of nocodazole (Wang, personal observation). Moreover, the sickness of Δcdc55 Δbub2 double mutants could not be rescued by Δswe1 deletion. Also, Δpph21 Δpph22 double mutants are capable of exiting mitosis without accumulating Swe1. Apparently, the premature mitotic exit phenotype in Δcdc55 mutants has nothing to do with Swe1 dependent Cdc28 phosphorylation. It is likely that PP2A has many substrates and the defects in the phosphorylation of different substrates contribute to the complex phenotype of Δcdc55 mutants. Swe1 protein accumulates in Δcdc55 mutants and deletion of PPH21 and PPH22 suppress Swe1 accumulation, suggesting that Cdc55 deregulates PP2A activity. But that is not the case for Cdc55 in the regulation of mitotic exit. First, Δpph21Δ pph22 double mutants exhibit benomyl sensitivity as do Δcdc55 mutants. Moreover, either Δcdc55 or Δpph21Δ pphh22 mutants suppress the AMN1 overexpression phenotype and both mutants exhibit nuclear localized Cdc14 in the presence of nocodazole, suggesting that mitotic exit pathways are hyperactive in both Δcdc55 and Δpph21 Δpph22 mutants. Consistently, deletion of TPD3, the A regulatory subunit of PP2A, also leads to less sensitivity to AMN1 overexpression. All these results indicate that both regulatory and catalytic subunits of PP2A are required for the negative regulation of mitotic exit pathways. It is the loss of function of PP2A in Δcdc55 mutants that causes premature mitotic exit. Bfa1 and Bub2 also negatively regulate mitotic exit by forming a complex with Tem1, a key player in the MEN pathway (Pereira et al., 2000). As Bfa1 is a phosphoprotein and its phosphorylation promotes mitotic exit (Hu et al., 2001), a reasonable model is that PP2A dephosphorylates Bfa1 to inhibit mitotic exit. However, our results argue against this model. The synthetic phenotype of Δcdc55 Δbfa1 and Δcdc55 Δbub2 double mutants indicates that
20 PP2A and Bfa1/Bub2 might act in concert and have additive effects on mitotic exit. Inactivation of both pathways results in deregulated Cdc14 localization and premature mitotic exit, which leads to the aberrant mitosis and sickness of the double mutants. Both FEAR and MEN pathways promote mitotic exit by stimulating Cdc14 release from the nucleolus. Our result indicates that defective FEAR pathway fails to block the mitotic exit completely in Δcdc55 mutants, as Δcdc55 Δslk19 double mutants are still able to rebud and release Cdc14 from the nucleolus in the presence of nocodazole. It is unlikely that PP2A negatively regulates mitotic exit by inhibiting FEAR pathway. MEN could be the target of PP2A based on the following observations. We have shown that AMN1 inhibits mitotic exit by binding to Tem1. Deletion of either CDC55 or PPH21 PPH22 suppresses AMN1 overexpression phenotype. Deletion of CDC55 also partially suppresses the temperature sensitivity of cdc5-1 and tem1-3, indicating that MEN is hyperactive in Δcdc55. However, cdc15-2 mutation completely suppresses the mitotic exit in Δcdc55 mutants when incubated at the restrictive temperature, suggesting that MEN is indispensable for the mitotic exit in Δcdc55 mutants. Importantly, Δcdc55 mutants show increased Tem1 protein phosphorylation. Therefore, the hyper active MEN in Δcdc55 mutants may result from the change of Tem1 protein modification. It is possible that a protein kinase phosphorylates Tem1 and activates MEN pathway, whereas PP2A dephosphorylates Tem1 and keeps it inactive. Defective PP2A will result in the increase of hyperphosphorylated Tem1, which promotes mitotic exit. In summary, we have identified a new layer of regulation of MEN, involving PP2A. In collaboration with Bfa1/Bub2 and Amn1, PP2A ensures cell cycle regulated localization of Cdc14, which is essential for a successful mitosis
21 Materials and methods: Yeast strains, growth and media: The genotype and sources of relevant yeast strains are listed in Table 1. All the strains listed are isogenic with W303 derived Y300 strain. All strains were constructed using standard genetic crosses. YYW28 was made by using a PCR based method (Longtine et al., 1998). To arrest yeast cells at G1 phase, 5μg/ml α-factor was added into mid-log cell cultures (OD 600 = 0.4) and the cultures were incubated for 2.5 hours. To release them into cell cycle, the cell cultures were centrifuged and washed once with H 2 O. Nocodazole was purchased from ICN and was used at 20μg/ml in a final concentration of 1% DMSO. Cytological techniques: Immunofluorescence straining was done after formaldehyde (3.7%) fixation for 15 minutes. Cells were treated with zymolase for 15 minutes and then stained with anti-ha antibody (1:100 Santa Cruz) overnight at 4 C on 14-well slides after methanol/acetone treatment. Afterwards, cells were stained with FITC-conjugated secondary antibody and DAPI and then visualized under immunofluorescence microscope (Zeiss). Protein techniques: 2 ml of cell culture was used to prepare protein samples for time course experiments. Cells were collected in tubes with screw caps after being centrifuged and 50μl of 20%TCA and glass beads were added. Cells were broken by using beads beater for 2 min. Protein was precipitated by centrifuge at 3,000 rpm for 2 min after glass beads were removed. Equal volume
22 (50μl) of 1M Tris-base and protein loading buffer were added. Dissolved protein samples were boiled for 5 min. 200 ml of cell culture (OD 600 =0.4) was used for Tem1 protein immuno-precipitation. Cells were collected by centrifugation and washed once with water and then resuspended in 0.5ml RIPA buffer supplied with protease and phosphatase inhibitors. Cells were broken with beads beater and the cell debris were removed after centrifuge at 14,000 rpm for 20 min at 4 C. 8μl of anti-myc antibody (from Convance) was added into the cell extract and the tube was shaken for 1.5 hr at 4 C. Then 50μl of anti-mouse IgG agarose beads was added and shaken for 1.5 hr. The beads were washed with 1X λ phosphatase buffer for three times and resuspended in 60μl 1X phosphatase buffer. The beads were then used for λ phosphatase treatment. In our experiment, NaF, β-glyceral phosphate, and Na 3 VO 4 were used as phosphatase inhibitor. Acknowledgments: We thank Drs Stephen Elledge, Angelica Amon, Jeff Bahant, Kyung Lee, Raymond Deshaies, Erfei Bi, James Broach, and John Pringle for plasmids and strains. We thank Dr. Daniel Burke for sharing the unpublished data. We thank Yanping Sun for preparing the media and buffers for the experiments. We also thank Dr. Stephen Elledge for his comments and Dr. LIoyd Epstein for thorough reading of this manuscript. This work was support by American Heart Association Scientist Development grant ( B) to Y.W. Y.W. was also supported by The Leukemia and Lymphoma Society Special Fellowship
23 Reference: Alexandru, G., Uhlmann, F., Mechtler, K., Poupart, M.A., and Nasmyth, K. (2001). Phosphorylation of the cohesin subunit Scc1 by Polo/Cdc5 kinase regulates sister chromatid separation in yeast. Cell 105, Alexandru, G., Zachariae, W., Schleiffer, A., and Nasmyth, K. (1999). Sister chromatid separation and chromosome re-duplication are regulated by different mechanisms in response to spindle damage. EMBO Journal 18, Bardin, A.J., Visintin, R., and Amon, A. (2000). A mechanism for coupling exit from mitosis to partitioning of the nucleus. Cell 102, Booher, R.N., Deshaies, R.J., and Kirschner, M.W. (1993). Properties of Saccharomyces cerevisiae wee1 and its differential regulation of p34cdc28 in response to G1 and G2 cyclins. EMBO J 12, Charles, J.F., Jaspersen, S.L., Tinker-Kulberg, R.L., Hwang, L., Szidon, A., and Morgan, D.O. (1998). The Polo-related kinase Cdc5 activates and is destroyed by the mitotic cyclin destruction machinery in S. cerevisiae. Current Biology 8, Evans, D.R., and Stark, M.J. (1997). Mutations in the Saccharomyces cerevisiae type 2A protein phosphatase catalytic subunit reveal roles in cell wall integrity, actin cytoskeleton organization and mitosis. Genetics 145, Geymonat, M., Spanos, A., Smith, S.J., Wheatley, E., Rittinger, K., Johnston, L.H., and Sedgwick, S.G. (2002). Control of mitotic exit in budding yeast. In vitro regulation of Tem1 GTPase by Bub2 and Bfa1. Journal of Biological Chemistry 277, Geymonat, M., Spanos, A., Walker, P.A., Johnston, L.H., and Sedgwick, S.G. (2003). In vitro regulation of budding yeast Bfa1/Bub2 GAP activity by Cdc5. J Biol Chem 278, Healy, A.M., Zolnierowicz, S., Stapleton, A.E., Goebl, M., DePaoli-Roach, A.A., and Pringle, J.R. (1991). CDC55, a Saccharomyces cerevisiae gene involved in cellular morphogenesis: identification, characterization, and homology to the B subunit of mammalian type 2A protein phosphatase. Molecular & Cellular Biology 11, Hoyt, M.A., Totis, L., and Roberts, B.T. (1991). S. cerevisiae genes required for cell cycle arrest in response to loss of microtubule function. Cell 66, Hu, F., Wang, Y., Liu, D., Li, Y., Qin, J., and Elledge, S.J. (2001). Regulation of the Bub2/Bfa1 GAP complex by Cdc5 and cell cycle checkpoints. Cell 107,
24 Hwang, L.H., Lau, L.F., Smith, D.L., Mistrot, C.A., Hardwick, K.G., Hwang, E.S., Amon, A., and Murray, A.W. (1998). Budding yeast Cdc20: a target of the spindle checkpoint. Science 279, Jaspersen, S.L., Charles, J.F., Tinker-Kulberg, R.L., and Morgan, D.O. (1998). A late mitotic regulatory network controlling cyclin destruction in Saccharomyces cerevisiae. Molecular Biology of the Cell 9, Komarnitsky, S.I., Chiang, Y.C., Luca, F.C., Chen, J., Toyn, J.H., Winey, M., Johnston, L.H., and Denis, C.L. (1998). DBF2 protein kinase binds to and acts through the cell cycle-regulated MOB1 protein. Molecular & Cellular Biology 18, Lee, S.E., Frenz, L.M., Wells, N.J., Johnson, A.L., and Johnston, L.H. (2001a). Order of function of the budding-yeast mitotic exit-network proteins Tem1, Cdc15, Mob1, Dbf2, and Cdc5. Current Biology 11, Lee, S.E., Jensen, S., Frenz, L.M., Johnson, A.L., Fesquet, D., and Johnston, L.H. (2001b). The Bub2-dependent mitotic pathway in yeast acts every cell cycle and regulates cytokinesis. Journal of Cell Science 114, Lengronne, A., and Schwob, E. (2002). The yeast CDK inhibitor Sic1 prevents genomic instability by promoting replication origin licensing in late G(1). Mol Cell 9, Li, R. (1999). Bifurcation of the mitotic checkpoint pathway in budding yeast. Proceedings of the National Academy of Sciences of the United States of America 96, Liu, H., Krizek, J., and Bretscher, A. (1992). Construction of a GAL1-regulated yeast cdna expression library and its application to the identification of genes whose overexpression causes lethality in yeast. Genetics 132, Longtine, M.S., McKenzie, A., 3rd, Demarini, D.J., Shah, N.G., Wach, A., Brachat, A., Philippsen, P., and Pringle, J.R. (1998). Additional modules for versatile and economical PCRbased gene deletion and modification in Saccharomyces cerevisiae. Yeast 14, Luca, F.C., and Winey, M. (1998). MOB1, an essential yeast gene required for completion of mitosis and maintenance of ploidy. Molecular Biology of the Cell 9, Mah, A.S., Jang, J., and Deshaies, R.J. (2001). Protein kinase Cdc15 activates the Dbf2-Mob1 kinase complex. Proceedings of the National Academy of Sciences of the United States of America 98, Michaelis, C., Ciosk, R., and Nasmyth, K. (1997). Cohesins: chromosomal proteins that prevent premature separation of sister chromatids. Cell 91, Millward, T.A., Zolnierowicz, S., and Hemmings, B.A. (1999). Regulation of protein kinase cascades by protein phosphatase 2A. Trends Biochem Sci 24,
25 Minshull, J., Straight, A., Rudner, A.D., Dernburg, A.F., Belmont, A., and Murray, A.W. (1996). Protein phosphatase 2A regulates MPF activity and sister chromatid cohesion in budding yeast. Current Biology 6, Morgan, D.O. (1999). Regulation of the APC and the exit from mitosis. Nature Cell Biology 1, E Noton, E., and Diffley, J.F. (2000). CDK inactivation is the only essential function of the APC/C and the mitotic exit network proteins for origin resetting during mitosis. Mol Cell 5, Pereira, G., Hofken, T., Grindlay, J., Manson, C., and Schiebel, E. (2000). The Bub2p spindle checkpoint links nuclear migration with mitotic exit. Molecular Cell 6, Ronne, H., Carlberg, M., Hu, G.Z., and Nehlin, J.O. (1991). Protein phosphatase 2A in Saccharomyces cerevisiae: effects on cell growth and bud morphogenesis. Mol Cell Biol 11, Schwab, M., Lutum, A.S., and Seufert, W. (1997). Yeast Hct1 is a regulator of Clb2 cyclin proteolysis. Cell 90, Shirayama, M., Matsui, Y., and Toh, E.A. (1994). The yeast TEM1 gene, which encodes a GTPbinding protein, is involved in termination of M phase. Mol Cell Biol 14, Shirayama, M., Zachariae, W., Ciosk, R., and Nasmyth, K. (1998). The Polo-like kinase Cdc5p and the WD-repeat protein Cdc20p/fizzy are regulators and substrates of the anaphase promoting complex in Saccharomyces cerevisiae. EMBO Journal 17, Shou, W., Seol, J.H., Shevchenko, A., Baskerville, C., Moazed, D., Chen, Z.W., Jang, J., Charbonneau, H., and Deshaies, R.J. (1999). Exit from mitosis is triggered by Tem1-dependent release of the protein phosphatase Cdc14 from nucleolar RENT complex. Cell 97, Stegmeier, F., Visintin, R., and Amon, A. (2002). Separase, polo kinase, the kinetochore protein Slk19, and Spo12 function in a network that controls Cdc14 localization during early anaphase. Cell 108, Vallen, E.A., Caviston, J., and Bi, E. (2000). Roles of Hof1p, Bni1p, Bnr1p, and myo1p in cytokinesis in Saccharomyces cerevisiae. Mol Biol Cell 11, Visintin, R., and Amon, A. (2001). Regulation of the mitotic exit protein kinases Cdc15 and Dbf2. Molecular Biology of the Cell 12, Visintin, R., Craig, K., Hwang, E.S., Prinz, S., Tyers, M., and Amon, A. (1998). The phosphatase Cdc14 triggers mitotic exit by reversal of Cdk-dependent phosphorylation. Molecular Cell 2,
26 Visintin, R., Hwang, E.S., and Amon, A. (1999). Cfi1 prevents premature exit from mitosis by anchoring Cdc14 phosphatase in the nucleolus.[comment]. Nature 398, Wang, Y., and Burke, D.J. (1997). Cdc55p, the B-type regulatory subunit of protein phosphatase 2A, has multiple functions in mitosis and is required for the kinetochore/spindle checkpoint in Saccharomyces cerevisiae. Molecular & Cellular Biology 17, Wang, Y., Shirogane, T., Liu, D., Harper, J.W., and Elledge, S.J. (2003). Exit from exit: resetting the cell cycle through Amn1 inhibition of G protein signaling. Cell 112, Yang, H., Jiang, W., Gentry, M., and Hallberg, R.L. (2000). Loss of a protein phosphatase 2A regulatory subunit (Cdc55p) elicits improper regulation of Swe1p degradation. Molecular & Cellular Biology 20,
27 Figure ligand: Figure 1. (A) Overexpression of PP2A components is lethal to mutants defective in mitotic exit. Saturated cell cultures were 10-time serial diluted and then spotted onto plates containing either glucose (Glu) or galactose (Gal). The plates were incubated at 25 C for 3 days. (B) and (C) Δcdc55 mutant suppresses the temperature sensitive phenotype of some mutants in the MEN pathway. The saturated cultures of strains with indicated genotype were 10-fold serial diluted and spotted onto YPD plates and incubated at indicated temperatures for 3 days. Figure 2. (A) Δcdc55 mutant cells exit mitosis in the presence of nocodazole. Wild type (Y300) and Δcdc55 mutant (YYW28) cells in mid-log phase were arrested at G1 phase with α- factor and then released into 30 C YPD medium containing 20μg/ml nocodazole after sonication. Cells were harvested at 30 minutes intervals for budding index. The top panel shows the percentile of cells with extra bud and the bottom panel shows cells after 240 min incubation in the presence of nocodazole. The arrow indicates the rebudded Δcdc55 mutant cell in the presence of nocodazole. (B) Top panel: Strain (Δcdc55 URA3-tetO-112 LEU2-tetR- GFP) was arrested at G1 phase and then released into YPD medium containing 20μg/ml nocodazole at 30 C. The picture was taken after 240 min incubation. The spots indicate GFP marked chromosome V. The bottom panel shows the frequency of reduplicated chromosome in wild type and Δcdc55 mutants. JBY583 (URA3-tetO-112 LEU2-tetR-GFP) and (Δcdc55 URA3-teto-112 LEU2-tetR-GFP) were arrested at G1 phase and then released into YPD medium containing 20μg/ml nocodazole. Cells were collected every 30 minutes and fixed for examination of the GFP signal. (C) The rebudding phenotype of Δcdc55 mutants is independent of Cdc28 Y19 phosphorylation. The top panel indicates the budding index of cells with
28 indicated genotype in YPD medium at 30 C. The bottom panel shows the percentage of rebudded cells in the presence of 20μg/ml of nocodazole. Strains (Y300, YYW28, , ) were treated as described in Figure 2A. Figure 3. (A) Δcdc55 mutant suppresses AMN1 overexpression phenotype. Strains (Y300, YFH240, YYW28, and YYW81) were transformed with either a CEN-URA3 vector or a P GAL - AMN1 plasmid. Saturated cultures of the transformants were 10-fold serial diluted and spotted onto plates containing glucose or galactose and incubated at 30 C for 2 days. (B) Mutations in catalytic subunits of PP2A result in benomyl sensitivity. Saturated cultures of strains with indicated genotype (Y300, YYW28, Y2475, and Y2762) were 10-fold serial diluted and spotted onto YPD and benomyl (15μg/ml) plates and incubated at 25 C for 3 days. (C) Δpph21 Δpph22 mutants suppress AMN1 overexpression phenotype. Y300 and Y2762 were transformed with either a CEN-URA3 vector or a P GAL -AMN1 plasmid. The transformants were treated as described in Figure 3A. (D) Δtpd3 mutant suppresses AMN1 overexpression phenotype (tpd3::kan) was transformed with either a vector or a P GAL -AMN1 plasmid and the growth of the transformants was examined as described. Figure 4. Cdc14 is released from the nucleolus in Δcdc55 mutants in the presence of nocodazole. (A) A1411 (CDC14-HA) and (Δcdc55 CDC14-HA) cells were arrested at G1 phase with α-factor and then released into YPD medium. Cells were collected every 20 minutes and fixed for Cdc14-HA staining. The budding index and the kinetics of Cdc14 release from the nucleolus are shown in the top and bottom panel respectively. (B) The same strains were arrested at G1 phase and then released into YPD medium containing 20μg/ml nocodazole at
29 30 C. The budding index and Cdc14 localization are shown. (C) The nuclear structure and Cdc14 localization of the 180 min samples in (B) are shown (top). The bottom of panel (C) shows the Net1 localization. G1 arrested NET1-myc and Δcdc55 NET1-myc cells were released into nocodazole medium. Cells were collected for Net1-myc staining. The nuclear structure and Net1 localization after 180 min incubation are shown. (D) Δcdc55 and Δpph21 Δpph22 mutants fail to prevent Cdc14 from releasing in the presence of nocodazole. Mid-log phase of A1411, and were incubated in the presence of 20μg/ml nocodazole at 30 C. Cells were collected after 3 hour incubation for Cdc14 staining. (E) Hof1 phosphorylation is more pronounced in nocodazole treated Δcdc55 mutants. G1 arrested (HOF1-HA) and (Δcdc55 HOF1-HA) cells were released into YPD medium containing 20μg/ml nocodazole at 30 C. Cells were collected every 30 minutes for protein preparation. Hof1-HA protein was examined by Western-Blot analysis after SDS-PAGE. Figure 5. CDC55 exhibits synthetic phenotype with BFA1 and BUB2. (A) G1 arrested BFA1- HA (YFH286) and Δcdc55 BFA1-HA ( ) cells were released into 30 C YPD medium. Cells were taken every 20 min to prepare protein extracts. The phosphorylation status of Bfa1 were determined by Western-Blot analysis with anti-ha antibodies. In the bottom panel, cdc14-1 BFA1-HA cells with a vector or a P GAL -CDC55 plasmid were grown to mid-log phase in the raffinose medium at 25 C followed by the following treatments: 1) to add galactose and incubate for 1 hour at 25 C; 2) to add galactose and incubate for 1 hour at 25 C and then shift to 34 C for 3 hours; 3) to shift to 34 C for 3 hours; 4) to shift to 34 C for 3 hours and then add galactose and incubate at 34 C for 1 hour. The phosphorylation of Bfa1 protein is shown after Western- Blotting with anti-ha antibody. (B) Δcdc55 Δbfa1 and Δcdc55 Δbub2 double mutants display
30 slow growth phenotype. Cells with indicated genotype were incubated at 30 C for 2 days. (C) The localization of Cdc14 in asynchronized Δcdc55 Δbfa1 ( ) double mutants. (D) Cell cycle regulated Cdc14 localization in Δcdc55 ( ), Δbub2 ( ) and Δcdc55 Δbub2 ( ) mutants. Cells were arrested at G1 phase with α-factor and then released into YPD medium at 30 C. 200mM hydroxyurea was added into the medium at 80 min when majority of the cells were large budded in order to block the release of Cdc14 from the nucleolus during the next cell cycle. Figure 6. Mitotic exit in Δcdc55 mutants depends on MEN pathway. (A) The localization of Cdc14 in Δslk19 and Δcdc55 Δslk19 mutants during unperturbed cell cycle progression. G1 arrested and were released into cell cycle at 30 and 200mM hydroxyurea was added into the medium at 80 min to block the next S phase. Cells were collected at 30 min interval for Cdc14 staining. (B) The left panel shows the budding morphology of cells with indicated genotypes. Strains with indicated genotypes were arrested at G1 and released into YPD medium containing 20μg/ml of nocodazole at 30 C for the determination of budding index. Right panel indicates the localization of Cdc14 in the presence of nocodazole. Strains with indicated genotype were released into nocodazole medium after G1 arrest. Cells were collected every 30 minutes and subjected to immunofluorescence staining for Cdc14-HA. The percentage of cells with nucleolar localized Cdc14 was determined over time. (C) cdc15-2 CDC14-HA and cdc15-2δ cdc55 CDC14-HA strains were arrested at G1 phase with α-factor and then released into YPD medium at 37 C. Budding index and the localization of Cdc14 were determined over time
31 Figure 7. PP2A regulates mitotic exit through Tem1 modification. (A) Δcdc55 deletion partially suppresses the mitotic exit defects of tem1-3 mutants. G1 arrested tem1-3, tem1-3 Δcdc55, cdc15-2, and cdc15-2 Δcdc55 mutants were released into 37 C YPD medium and the budding morphology was examined over time. The percentage of rebudded cells is shown. (B) Tem1 is a phosphoprotein. Tem1-myc fusion protein was immunoprecipitated from the cell lysate of cdc14-1 TEM1-myc incubated at 36 C for 2 hr. The precipitates were incubated with λ phosphatase at 30 C for 30 min in the presence or absence of phosphatase inhibitors. Proteins were separated with 10% SDS-PAGE and then subjected to Western-Blotting with anti-myc antibody. (C) Δcdc55 mutation leads to hyperphosphorylation of Tem (TEM1-13myc) and (Δcdc55 TEM1-13myc) were arrested at G1 phase and then released into 30 C YPD medium containing 20μg/ml nocodazole. Protein samples were prepared every 20 min with TCA method. Proteins were separated by SDS-PAGE followed by Western-Blot analysis. Top panel shows the percentage of large budded cells and the bottom panel shows the modification of Tem1 protein. (D) Protein samples from 0, 80 and 140 min time points in Figure 7C were loaded side by side and the band shift of Tem1 protein is shown after Western Blot analysis
32 Table 1. Strains used in this study Strains Relevant Genotype Source Y300 Mata ura3-1 his3-11, 15 leu2-3, 112 trp1-1 ade2-1 can1-100 Elledge lab JBY517 Mata cdc5-1 J. Bachant Mata mob1-77 This study Mata tem1-3 This study Mata tem1-3 Δcdc55::his5 + This study Mata cdc15-2 This study Mata cdc15-2 Δcdc55::his5 + This study Mata cdc5-1 Δcdc55:his5 + This study YYW28 Mata Δcdc55::his5 + This study Mata Δcdc55::his5 + Δcdc28::URA3 CDC28F19-TRP1 This study Mata Δcdc28::URA3 CDC28F19-TRP1 This study Mata Δcdc55::his5 + URA3-tetO-112 LEU2-tetR-GFP This study JBY583 Mata tetr-gfp-leu2, teto-ura3 in Y300 J. Bachant YFH240 Mata Δbfa1::his5 + Elledge lab YYW81 Mata Δmad1::HIS3 in Y300 This study Y2475 Mata Δpph21::HIS3 J. Broach Y2762 Mata Δpph21::LEU2 Δpph22::HIS3 J. Broach Mata Δtpd3::Kan This study YYW1 Mata Δbub2::URA3 This study Mata Δbfa1::his5 + Δcdc55::his5 + This study Mata Δbub2::URA3 Δcdc55:his5 + This study Mata Δbfa1::his5 + Δcdc55::his5 + CDC14-HA This study A1411 Mata CDC14-HA A. Amon Mata Δcdc55::his5 + CDC14-HA This study YYW68-1 Mata Δslk19::TRP1 This study Mata Δcdc55::his5 + Δslk19::TRP1 This study Mata Δcdc55::his5 + Δslk19::TRP1 CDC14-HA This study Mata cdc15-2 CDC14-HA This study Mata cdc15-2 Δcdc55::his5 + CDC14-HA This study Mata Δpph21::LEU2 Δpph22::HIS3 CDC14-HA This study Mata Δbub2::URA3 CDC14-HA This study Mata Δbub2::URA3 Δcdc55::his5 + Δswe1::LEU2 CDC14-HA This study Mata NET1-TEV-myc9-his5 + This Study Mata cdc55:his5 + NET1-TEV-myc9-his5 + This Study Mata HOF-HA-his5 + This Study Mata Δcdc55::his5 + HOF-HA-his5 + This Study YFH286 Mata BFA-HA-TRP1 Elledge lab Mata Δcdc55::his5 + BFA1-HA3-TRP1 This Study Mata TEM1-13myc-kan This Study Mata Δcdc55::his5 + TEM1-13myc-Kan This Study

33
34
35
36
37
38
39
7.06 Problem Set #4, Spring 2005
 7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
7.06 Problem Set #4, Spring 2005 1. You re doing a mutant hunt in S. cerevisiae (budding yeast), looking for temperaturesensitive mutants that are defective in the cell cycle. You discover a mutant strain
Department of Pharmacology, UT Southwestern Medical Center, 5323 Harry Hines Blvd., Dallas, TX
 [Frontiers in Bioscience 8, d1275-1287, September 1, 2003] REGULATION OF CDC14: PATHWAYS AND CHECKPOINTS OF MITOTIC EXIT Joshua Bembenek and Hongtao Yu Department of Pharmacology, UT Southwestern Medical
[Frontiers in Bioscience 8, d1275-1287, September 1, 2003] REGULATION OF CDC14: PATHWAYS AND CHECKPOINTS OF MITOTIC EXIT Joshua Bembenek and Hongtao Yu Department of Pharmacology, UT Southwestern Medical
Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter
 Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter 9/10/2008 1 Learning Objectives Explain why a cell cycle was selected for during evolution
Plant Molecular and Cellular Biology Lecture 8: Mechanisms of Cell Cycle Control and DNA Synthesis Gary Peter 9/10/2008 1 Learning Objectives Explain why a cell cycle was selected for during evolution
Lecture 10: Cyclins, cyclin kinases and cell division
 Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Chem*3560 Lecture 10: Cyclins, cyclin kinases and cell division The eukaryotic cell cycle Actively growing mammalian cells divide roughly every 24 hours, and follow a precise sequence of events know as
Analysis and Simulation of Biological Systems
 Analysis and Simulation of Biological Systems Dr. Carlo Cosentino School of Computer and Biomedical Engineering Department of Experimental and Clinical Medicine Università degli Studi Magna Graecia Catanzaro,
Analysis and Simulation of Biological Systems Dr. Carlo Cosentino School of Computer and Biomedical Engineering Department of Experimental and Clinical Medicine Università degli Studi Magna Graecia Catanzaro,
Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells
 Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells Understand how a simple biochemical oscillator can drive the
Life Sciences 1a: Section 3B. The cell division cycle Objectives Understand the challenges to producing genetically identical daughter cells Understand how a simple biochemical oscillator can drive the
16 The Cell Cycle. Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization
 The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
The Cell Cycle 16 The Cell Cycle Chapter Outline The Eukaryotic Cell Cycle Regulators of Cell Cycle Progression The Events of M Phase Meiosis and Fertilization Introduction Self-reproduction is perhaps
A Mathematical Model of Mitotic Exit in Budding Yeast: The Role of Polo Kinase
 A Mathematical Model of Mitotic Exit in Budding Yeast: The Role of Polo Kinase Baris Hancioglu*, John J. Tyson Department of Biological Sciences, Virginia Polytechnic Institute and State University, Blacksburg,
A Mathematical Model of Mitotic Exit in Budding Yeast: The Role of Polo Kinase Baris Hancioglu*, John J. Tyson Department of Biological Sciences, Virginia Polytechnic Institute and State University, Blacksburg,
A redundant function for the N-terminal tail of Ndc80 in kinetochore-microtubule interaction in Saccharomyces cerevisiae.
 Genetics: Published Articles Ahead of Print, published on July 30, 2012 as 10.1534/genetics.112.143818 A redundant function for the N-terminal tail of Ndc80 in kinetochore-microtubule interaction in Saccharomyces
Genetics: Published Articles Ahead of Print, published on July 30, 2012 as 10.1534/genetics.112.143818 A redundant function for the N-terminal tail of Ndc80 in kinetochore-microtubule interaction in Saccharomyces
Regulation of the Cell Cycle by Protein Phosphatase 2A in Saccharomyces cerevisiae
 MICROBIOLOGY AND MOLECULAR BIOLOGY REVIEWS, June 2006, p. 440 449 Vol. 70, No. 2 1092-2172/06/$08.00 0 doi:10.1128/mmbr.00049-05 Copyright 2006, American Society for Microbiology. All Rights Reserved.
MICROBIOLOGY AND MOLECULAR BIOLOGY REVIEWS, June 2006, p. 440 449 Vol. 70, No. 2 1092-2172/06/$08.00 0 doi:10.1128/mmbr.00049-05 Copyright 2006, American Society for Microbiology. All Rights Reserved.
Role of the mitotic cyclin Clb2 in mitotic regulation
 University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange Masters Theses Graduate School 8-2010 Role of the mitotic cyclin Clb2 in mitotic regulation Dustin K. Stutts dkimbrou@utk.edu
University of Tennessee, Knoxville Trace: Tennessee Research and Creative Exchange Masters Theses Graduate School 8-2010 Role of the mitotic cyclin Clb2 in mitotic regulation Dustin K. Stutts dkimbrou@utk.edu
Overview of the cell cycle
 Chapter 2 Overview of the cell cycle 2.1 The organisation of cell cycle in eukaryotes During the cell cycle, the typical eukaryotic cell goes through a series of well defined phases, to divide into two
Chapter 2 Overview of the cell cycle 2.1 The organisation of cell cycle in eukaryotes During the cell cycle, the typical eukaryotic cell goes through a series of well defined phases, to divide into two
Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc
 OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
OPTIMIZATION OF IMMUNOBLOT PROTOCOL 121 Optimization of Immunoblot Protocol for Use with a Yeast Strain Containing the CDC7 Gene Tagged with myc Jacqueline Bjornton and John Wheeler Faculty Sponsor: Anne
Three different fusions led to three basic ideas: 1) If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes
 If one fuses a cell in mitosis with a cell in any other stage of the cell cycle, the chromosomes") Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Section Notes The cell division cycle presents an interesting system to study because growth and division must be carefully coordinated. For many cells it is important that it reaches the correct size
Dr. Fred Cross, Rockefeller (KITP Bio Networks 3/26/2003) 1
 1") Outline Cell growth as the driver for cell cycle (in microbes): coordination of growth and division A basic principle organizing cell cycle control: why cyclin-dependent kinase activity must oscillate
Outline Cell growth as the driver for cell cycle (in microbes): coordination of growth and division A basic principle organizing cell cycle control: why cyclin-dependent kinase activity must oscillate
12/5/2014. The cell cycle and cell death. The cell cycle: cells duplicate their contents and divide
 The cell cycle and cell death The cell cycle: cells duplicate their contents and divide 1 The cell cycle may be divided into 4 phases Eucaryotic cell division: Mitosis (nuclear division) Cytokinesis (cell
The cell cycle and cell death The cell cycle: cells duplicate their contents and divide 1 The cell cycle may be divided into 4 phases Eucaryotic cell division: Mitosis (nuclear division) Cytokinesis (cell
Lecture Series 5 Cell Cycle & Cell Division
 Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Division Read Chapter 19 pages 651-663 663 only (Benefits of Sex & Meiosis sections these are in Chapter
Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Division Read Chapter 19 pages 651-663 663 only (Benefits of Sex & Meiosis sections these are in Chapter
1- Below is a list of cell cycle phases matched with specific processes. Choose the correct pairing:
 Name: NetID: Exam 4 - Version 2 November 13, 2018 Dr. A. Pimentel Instructions: 1- Select the BEST answer for each question 2- Use pencil to mark your responses in the answer sheet. 3- You can mark your
Name: NetID: Exam 4 - Version 2 November 13, 2018 Dr. A. Pimentel Instructions: 1- Select the BEST answer for each question 2- Use pencil to mark your responses in the answer sheet. 3- You can mark your
Reading Assignments. A. Systems of Cell Division. Lecture Series 5 Cell Cycle & Cell Division
 Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
Lecture Series 5 Cell Cycle & Cell Division
 Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
Lecture Series 5 Cell Cycle & Cell Division Reading Assignments Read Chapter 18 Cell Cycle & Cell Death Read Chapter 19 Cell Division Read Chapter 20 pages 659-672 672 only (Benefits of Sex & Meiosis sections)
The cell cycle entails an ordered series of macromolecular
 21 REGULATING THE EUKARYOTIC CELL CYCLE This cultured rat kidney cell in metaphase shows condensed chromosomes (blue), microtubules of the spindle apparatus (red), and the inner nuclear envelope protein
21 REGULATING THE EUKARYOTIC CELL CYCLE This cultured rat kidney cell in metaphase shows condensed chromosomes (blue), microtubules of the spindle apparatus (red), and the inner nuclear envelope protein
Essential Knowledge: In eukaryotes, heritable information is passed to the next generation via processes that include the cell cycle and mitosis OR
 Essential Knowledge: In eukaryotes, heritable information is passed to the next generation via processes that include the cell cycle and mitosis OR meiosis plus fertilization Objective: You will be able
Essential Knowledge: In eukaryotes, heritable information is passed to the next generation via processes that include the cell cycle and mitosis OR meiosis plus fertilization Objective: You will be able
Biology: Life on Earth
 Biology: Life on Earth Eighth Edition Lecture for Chapter 11 The Continuity of Life: Cellular Reproduction Cellular Reproduction Intracellular activity between one cell division to the next is the cell
Biology: Life on Earth Eighth Edition Lecture for Chapter 11 The Continuity of Life: Cellular Reproduction Cellular Reproduction Intracellular activity between one cell division to the next is the cell
The Cell Cycle/Le Cycle cellulaire SMC6052/BIM6028 IRCM
 The Cell Cycle/Le Cycle cellulaire SMC6052/BIM6028 IRCM 1 février 2018 Benjamin H. Kwok, Ph.D. Chercheur principal, Institut de recherche en immunologie et en cancérologie Professeur sous octroi agrégé,
The Cell Cycle/Le Cycle cellulaire SMC6052/BIM6028 IRCM 1 février 2018 Benjamin H. Kwok, Ph.D. Chercheur principal, Institut de recherche en immunologie et en cancérologie Professeur sous octroi agrégé,
The FEAR network. Citation. As Published Publisher. Version
 The FEAR network The MIT Faculty has made this article openly available. lease share how this access benefits you. Your story matters. Citation As ublished ublisher Rock, Jeremy M., and Angelika Amon.
The FEAR network The MIT Faculty has made this article openly available. lease share how this access benefits you. Your story matters. Citation As ublished ublisher Rock, Jeremy M., and Angelika Amon.
3.a.2- Cell Cycle and Meiosis
 Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. 3.a.2- Cell Cycle and Meiosis EU 3.A: Heritable information provides for continuity of life.
Big Idea 3: Living systems store, retrieve, transmit and respond to information essential to life processes. 3.a.2- Cell Cycle and Meiosis EU 3.A: Heritable information provides for continuity of life.
AP Biology Unit 6 Practice Test 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8
 AP Biology Unit 6 Practice Test Name: 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8 picograms of DNA per nucleus. How many picograms
AP Biology Unit 6 Practice Test Name: 1. A group of cells is assayed for DNA content immediately following mitosis and is found to have an average of 8 picograms of DNA per nucleus. How many picograms
Research Paper 497. Address: Departments of Physiology and Biochemistry & Biophysics, University of California, San Francisco, California 94143, USA.
 Research Paper 497 The Polo-related kinase activates and is destroyed by the mitotic cyclin destruction machinery in S. cerevisiae Julia F. Charles, Sue. Jaspersen, Rachel. Tinker-Kulberg, ena Hwang, Alex
Research Paper 497 The Polo-related kinase activates and is destroyed by the mitotic cyclin destruction machinery in S. cerevisiae Julia F. Charles, Sue. Jaspersen, Rachel. Tinker-Kulberg, ena Hwang, Alex
Study Guide 11 & 12 MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question.
 Study Guide 11 & 12 MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) The receptors for a group of signaling molecules known as growth factors are
Study Guide 11 & 12 MULTIPLE CHOICE. Choose the one alternative that best completes the statement or answers the question. 1) The receptors for a group of signaling molecules known as growth factors are
CHAPTER 12 - THE CELL CYCLE (pgs )
") CHAPTER 12 - THE CELL CYCLE (pgs. 228-245) CHAPTER SEVEN TARGETS I. Describe the importance of mitosis in single-celled and multi-cellular organisms. II. Explain the organization of DNA molecules and their
CHAPTER 12 - THE CELL CYCLE (pgs. 228-245) CHAPTER SEVEN TARGETS I. Describe the importance of mitosis in single-celled and multi-cellular organisms. II. Explain the organization of DNA molecules and their
ACCELERATE ITS BIOCHEMICAL PROCESSES WHICH WERE SLOWED DOWN BY MITOSIS. THE LENGTH OF THE G1 PHASE CREATES THE DIFFERENCE BETWEEN FAST DIVIDING
 CHAPTER 1: OVERVIEW OF THE CELL CYCLE THE THREE STAGES OF INTERPHASE: INTERPHASE BEFORE A CELL CAN ENTER CELL DIVISION, IT NEEDS TO PREPARE ITSELF BY REPLICATING ITS GENETIC INFORMATION AND ALL OF THE
CHAPTER 1: OVERVIEW OF THE CELL CYCLE THE THREE STAGES OF INTERPHASE: INTERPHASE BEFORE A CELL CAN ENTER CELL DIVISION, IT NEEDS TO PREPARE ITSELF BY REPLICATING ITS GENETIC INFORMATION AND ALL OF THE
What happens to the replicated chromosomes? depends on the goal of the division
 Segregating the replicated chromosomes What happens to the replicated chromosomes? depends on the goal of the division - to make more vegetative cells: mitosis daughter cells chromosome set should be identical
Segregating the replicated chromosomes What happens to the replicated chromosomes? depends on the goal of the division - to make more vegetative cells: mitosis daughter cells chromosome set should be identical
Cell cycle regulation in the budding yeast
 Cell cycle regulation in the budding yeast Bởi: TS. Nguyen Cuong Introduction The cell cycle is the sequence of events by which a growing cell duplicates all its components and then divides into two daughter
Cell cycle regulation in the budding yeast Bởi: TS. Nguyen Cuong Introduction The cell cycle is the sequence of events by which a growing cell duplicates all its components and then divides into two daughter
Table S1. Aspergillus nidulans strains used in this study Strain Genotype Derivation
 Supplemental Material De Souza et al., 211 Table S1. Aspergillus nidulans strains used in this study Strain Genotype Derivation CDS295 pyrg89; pyroa4; pyrg Af ::son promotor::gfp-son nup98/nup96 ; chaa1
Supplemental Material De Souza et al., 211 Table S1. Aspergillus nidulans strains used in this study Strain Genotype Derivation CDS295 pyrg89; pyroa4; pyrg Af ::son promotor::gfp-son nup98/nup96 ; chaa1
Sic1 as a timer of Clb cyclin waves in the yeast cell cycle design principle of not just an inhibitor
 REVIEW ARTICLE Sic1 as a timer of Clb cyclin waves in the yeast cell cycle design principle of not just an inhibitor Matteo Barberis 1,2 1 Institute for Biology, Theoretical Biophysics, Humboldt University
REVIEW ARTICLE Sic1 as a timer of Clb cyclin waves in the yeast cell cycle design principle of not just an inhibitor Matteo Barberis 1,2 1 Institute for Biology, Theoretical Biophysics, Humboldt University
A Late Mitotic Regulatory Network Controlling Cyclin Destruction in Saccharomyces cerevisiae
 Molecular Biology of the Cell Vol. 9, 2803 2817, October 1998 A Late Mitotic Regulatory Network Controlling Cyclin Destruction in Saccharomyces cerevisiae Sue L. Jaspersen, Julia F. Charles, Rachel L.
Molecular Biology of the Cell Vol. 9, 2803 2817, October 1998 A Late Mitotic Regulatory Network Controlling Cyclin Destruction in Saccharomyces cerevisiae Sue L. Jaspersen, Julia F. Charles, Rachel L.
Control of the mitotic exit network during meiosis
 Control of the mitotic exit network during meiosis The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published Publisher
Control of the mitotic exit network during meiosis The MIT Faculty has made this article openly available. Please share how this access benefits you. Your story matters. Citation As Published Publisher
BIOLOGY. Chapter 10 CELL REPRODUCTION PowerPoint Image Slideshow
 BIOLOGY Chapter 10 CELL REPRODUCTION PowerPoint Image Slideshow FIGURE 10.1 A sea urchin begins life as a single cell that (a) divides to form two cells, visible by scanning electron microscopy. After
BIOLOGY Chapter 10 CELL REPRODUCTION PowerPoint Image Slideshow FIGURE 10.1 A sea urchin begins life as a single cell that (a) divides to form two cells, visible by scanning electron microscopy. After
Spindle Position Is Coordinated with Cell-Cycle Progression through Establishment of Mitotic Exit-Activating and -Inhibitory Zones
 Article Spindle Position Is Coordinated with Cell-Cycle Progression through Establishment of Mitotic Exit-Activating and -Inhibitory Zones Leon Y. Chan 1 and Angelika Amon 1, * 1 David H. Koch Institute
Article Spindle Position Is Coordinated with Cell-Cycle Progression through Establishment of Mitotic Exit-Activating and -Inhibitory Zones Leon Y. Chan 1 and Angelika Amon 1, * 1 David H. Koch Institute
Regulation of Protein Kinase Dbf2 in Mitotic Exit
 Regulation of Protein Kinase Dbf2 in Mitotic Exit Thesis by Angie S. Mah In Partial Fulfillment of the Requirements For the Degree of Doctor of Philosophy California Institute of Technology Pasadena, California
Regulation of Protein Kinase Dbf2 in Mitotic Exit Thesis by Angie S. Mah In Partial Fulfillment of the Requirements For the Degree of Doctor of Philosophy California Institute of Technology Pasadena, California
CELL CYCLE AND GROWTH REGULATION
 CELL CYCLE AND GROWTH REGULATION The cell cycle is the set of stages through which a cell progresses from one division to the next. Interphase is the period between mitotic cell divisions; divided into
CELL CYCLE AND GROWTH REGULATION The cell cycle is the set of stages through which a cell progresses from one division to the next. Interphase is the period between mitotic cell divisions; divided into
A simple model for the eukaryotic cell cycle. Andrea Ciliberto
 A simple model for the eukaryotic cell cycle Andrea Ciliberto The cell division cycle G1 cell division Start S (DNA Replication) Finish M (mitosis) G2/M G2 Kohn, Mol. Biol. Cell., 1999 How did we get to
A simple model for the eukaryotic cell cycle Andrea Ciliberto The cell division cycle G1 cell division Start S (DNA Replication) Finish M (mitosis) G2/M G2 Kohn, Mol. Biol. Cell., 1999 How did we get to
AP Biology - Cell cycle / division
 AP Biology - Cell cycle / division Quiz Directions 1. During which stage does DNA replication occur? A. Prophase B. Metaphase C. Anaphase D. none of these 2. At what phase in the cell cycle does DNA replication
AP Biology - Cell cycle / division Quiz Directions 1. During which stage does DNA replication occur? A. Prophase B. Metaphase C. Anaphase D. none of these 2. At what phase in the cell cycle does DNA replication
AP Biology Fall Semester Set 1
 1. During which stage does DNA replication occur? A. Prophase B. Metaphase C. Anaphase D. none of these 2. At what phase in the cell cycle does DNA replication occur? A. G1 B. S C. G2 D. M 3. Which of
1. During which stage does DNA replication occur? A. Prophase B. Metaphase C. Anaphase D. none of these 2. At what phase in the cell cycle does DNA replication occur? A. G1 B. S C. G2 D. M 3. Which of
A Few Terms: When and where do you want your cells to divide?
 Today: - Lab 4 Debrief - Mitosis - Lunch -Meiosis Other: Blood Drive Today! TIME: 11:00am 1:00pm + 2:00pm 5:00pm PLACE: Baxter Events Center Thinking About Mitosis When and where do you want your cells
Today: - Lab 4 Debrief - Mitosis - Lunch -Meiosis Other: Blood Drive Today! TIME: 11:00am 1:00pm + 2:00pm 5:00pm PLACE: Baxter Events Center Thinking About Mitosis When and where do you want your cells
Describe the process of cell division in prokaryotic cells. The Cell Cycle
 The Cell Cycle Objective # 1 In this topic we will examine the cell cycle, the series of changes that a cell goes through from one division to the next. We will pay particular attention to how the genetic
The Cell Cycle Objective # 1 In this topic we will examine the cell cycle, the series of changes that a cell goes through from one division to the next. We will pay particular attention to how the genetic
Dr. Mahmood S. Choudhery, PhD, Postdoc (USA) Assistant Professor Tissue Engineering & Regenerative Medicine King Edward Medical University
 Assistant Professor Tissue Engineering & Regenerative Medicine King Edward Medical University") CELL DIVISION Dr. Mahmood S. Choudhery, PhD, Postdoc (USA) Assistant Professor Tissue Engineering & Regenerative Medicine King Edward Medical University Cell Division The key roles of cell division Unicellular
CELL DIVISION Dr. Mahmood S. Choudhery, PhD, Postdoc (USA) Assistant Professor Tissue Engineering & Regenerative Medicine King Edward Medical University Cell Division The key roles of cell division Unicellular
cellular division cell division cell cycle cell cycle kinases chapter 18-19
 cellular division chapter 18-19 cell division when? growth replacement of older cells production of specialized cells asexual reproduction sexual reproduction production of gametes prokaryotic binary fission
cellular division chapter 18-19 cell division when? growth replacement of older cells production of specialized cells asexual reproduction sexual reproduction production of gametes prokaryotic binary fission
Edinburgh Research Explorer
 Edinburgh Research Explorer Cdc55 coordinates spindle assembly and chromosome disjunction during meiosis Citation for published version: Bizzari, F & Marston, AL 2011, 'Cdc55 coordinates spindle assembly
Edinburgh Research Explorer Cdc55 coordinates spindle assembly and chromosome disjunction during meiosis Citation for published version: Bizzari, F & Marston, AL 2011, 'Cdc55 coordinates spindle assembly
10 CELL DIVISION AND MITOSIS
 10 CELL DIVISION AND MITOSIS Chapter Outline Why It Matters 10.1 THE CYCLE OF CELL GROWTH AND DIVISION: OVERVIEW The products of mitosis are genetic duplicates of the dividing cell Chromosomes are the
10 CELL DIVISION AND MITOSIS Chapter Outline Why It Matters 10.1 THE CYCLE OF CELL GROWTH AND DIVISION: OVERVIEW The products of mitosis are genetic duplicates of the dividing cell Chromosomes are the
Cdk-counteracting phosphatases unlock mitotic exit Ethel Queralt 1 and Frank Uhlmann 2
 Available online at www.sciencedirect.com Cdk-counteracting phosphatases unlock mitotic exit Ethel Queralt 1 and Frank Uhlmann 2 Entry into mitosis of the eukaryotic cell cycle is driven by rising cyclin-dependent
Available online at www.sciencedirect.com Cdk-counteracting phosphatases unlock mitotic exit Ethel Queralt 1 and Frank Uhlmann 2 Entry into mitosis of the eukaryotic cell cycle is driven by rising cyclin-dependent
Zool 3200: Cell Biology Exam 5 4/27/15
 Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Name: Trask Zool 3200: Cell Biology Exam 5 4/27/15 Answer each of the following short answer questions in the space provided, giving explanations when asked to do so. Circle the correct answer or answers
Fertilization of sperm and egg produces offspring
 In sexual reproduction Fertilization of sperm and egg produces offspring In asexual reproduction Offspring are produced by a single parent, without the participation of sperm and egg CONNECTIONS BETWEEN
In sexual reproduction Fertilization of sperm and egg produces offspring In asexual reproduction Offspring are produced by a single parent, without the participation of sperm and egg CONNECTIONS BETWEEN
Honors Biology Test Chapter 8 Mitosis and Meiosis
 Honors Biology Test Chapter 8 Mitosis and Meiosis 1. In mitosis, if a parent cell has 16 chromosomes, each daughter cell will have how many chromosomes? a. 64 b. 32 c. 16 d. 8 e. 4 2. Chromatids that are
Honors Biology Test Chapter 8 Mitosis and Meiosis 1. In mitosis, if a parent cell has 16 chromosomes, each daughter cell will have how many chromosomes? a. 64 b. 32 c. 16 d. 8 e. 4 2. Chromatids that are
Name 8 Cell Cycle and Meiosis Test Date Study Guide You must know: The structure of the replicated chromosome. The stages of mitosis.
 Name 8 Cell Cycle and Meiosis Test Date Study Guide You must know: The structure of the replicated chromosome. The stages of mitosis. The role of kinases and cyclin in the regulation of the cell cycle.
Name 8 Cell Cycle and Meiosis Test Date Study Guide You must know: The structure of the replicated chromosome. The stages of mitosis. The role of kinases and cyclin in the regulation of the cell cycle.
Meiosis. Bởi: OpenStaxCollege
 Meiosis Bởi: OpenStaxCollege Sexual reproduction requires fertilization, a union of two cells from two individual organisms. If those two cells each contain one set of chromosomes, then the resulting cell
Meiosis Bởi: OpenStaxCollege Sexual reproduction requires fertilization, a union of two cells from two individual organisms. If those two cells each contain one set of chromosomes, then the resulting cell
Name Chapter 10: Chromosomes, Mitosis, and Meiosis Mrs. Laux Take home test #7 DUE: MONDAY, NOVEMBER 16, 2009 MULTIPLE CHOICE QUESTIONS
 MULTIPLE CHOICE QUESTIONS 1. A bacterial chromosome consists of: A. a linear DNA molecule many times larger than the cell. B. a circular DNA molecule many times larger than the cell. C. a circular DNA
MULTIPLE CHOICE QUESTIONS 1. A bacterial chromosome consists of: A. a linear DNA molecule many times larger than the cell. B. a circular DNA molecule many times larger than the cell. C. a circular DNA
Chapter 12: The Cell Cycle. 2. What is the meaning of genome? Compare your genome to that of a prokaryotic cell.
 Name: AP Bio Chapter 12: The Cell Cycle 12.1 Cell division results in genetically identical daughter cells 1. What is meant by the cell cycle? 2. What is the meaning of genome? Compare your genome to that
Name: AP Bio Chapter 12: The Cell Cycle 12.1 Cell division results in genetically identical daughter cells 1. What is meant by the cell cycle? 2. What is the meaning of genome? Compare your genome to that
Cyclin Dependent Kinases and Cell Cycle Control
 Bioscience Reports, Vol. 22, Nos. 5 and 6, October and December 2002 ( 2002) NOBEL LECTURE 9 DECEMBER, 2001 Cyclin Dependent Kinases and Cell Cycle Control Paul M. Nurse The discovery of major regulators
Bioscience Reports, Vol. 22, Nos. 5 and 6, October and December 2002 ( 2002) NOBEL LECTURE 9 DECEMBER, 2001 Cyclin Dependent Kinases and Cell Cycle Control Paul M. Nurse The discovery of major regulators
2. Which of the following are NOT prokaryotes? A) eubacteria B) archaea C) viruses D) ancient bacteria
 eubacteria B) archaea C) viruses D) ancient bacteria") 1. Which of the following statements is FALSE? A) Errors in chromosome separation are rarely a problem for an organism. B) Errors in chromosome separation can result in a miscarriage. C) Errors in chromosome
1. Which of the following statements is FALSE? A) Errors in chromosome separation are rarely a problem for an organism. B) Errors in chromosome separation can result in a miscarriage. C) Errors in chromosome
Human Biology Chapter 13.4: Meiosis and Genetic Variation
 OpenStax-CNX module: m58013 1 Human Biology Chapter 13.4: Meiosis and Genetic Variation Willy Cushwa Based on Meiosis by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons
OpenStax-CNX module: m58013 1 Human Biology Chapter 13.4: Meiosis and Genetic Variation Willy Cushwa Based on Meiosis by OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons
Unit 6 Test: The Cell Cycle
 Name Date Class Mrs. Knight Biology EHS Unit 6 Test: The Cell Cycle 1. What are the four main stages of the cell cycle (correct order)? A. G 1, S, G 0, M C. G 2, S, G 1, M B. G 1, S, G 2, M D. M, G 2,
Name Date Class Mrs. Knight Biology EHS Unit 6 Test: The Cell Cycle 1. What are the four main stages of the cell cycle (correct order)? A. G 1, S, G 0, M C. G 2, S, G 1, M B. G 1, S, G 2, M D. M, G 2,
Meiosis * OpenStax. This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0.
 OpenStax-CNX module: m45466 1 Meiosis * OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section, you will be able to: Abstract
OpenStax-CNX module: m45466 1 Meiosis * OpenStax This work is produced by OpenStax-CNX and licensed under the Creative Commons Attribution License 3.0 By the end of this section, you will be able to: Abstract
Mad2 transiently associates with an APC p55cdc complex during mitosis
 Proc. Natl. Acad. Sci. USA Vol. 95, pp. 11193 11198, September 1998 Cell Biology Mad2 transiently associates with an APC p55cdc complex during mitosis KATJA WASSMANN AND ROBERT BENEZRA* Cell Biology and
Proc. Natl. Acad. Sci. USA Vol. 95, pp. 11193 11198, September 1998 Cell Biology Mad2 transiently associates with an APC p55cdc complex during mitosis KATJA WASSMANN AND ROBERT BENEZRA* Cell Biology and
Introduction: The Cell Cycle and Mitosis
 Contents 1 Introduction: The Cell Cycle and Mitosis 2 Mitosis Review Introduction: The Cell Cycle and Mitosis The cell cycle refers to the a series of events that describe the metabolic processes of growth
Contents 1 Introduction: The Cell Cycle and Mitosis 2 Mitosis Review Introduction: The Cell Cycle and Mitosis The cell cycle refers to the a series of events that describe the metabolic processes of growth
Edinburgh Research Explorer
 Edinburgh Research Explorer Meiosis Citation for published version: Marston, AL 2009, 'Meiosis: DDK is not just for replication' Current biology : CB, vol. 19, no. 2, pp. R74-6. DOI: 10.1016/j.cub.2008.12.003
Edinburgh Research Explorer Meiosis Citation for published version: Marston, AL 2009, 'Meiosis: DDK is not just for replication' Current biology : CB, vol. 19, no. 2, pp. R74-6. DOI: 10.1016/j.cub.2008.12.003
Sexual Reproduction and Meiosis. Chapter 11
 Sexual Reproduction and Meiosis Chapter 11 1 Sexual life cycle Made up of meiosis and fertilization Diploid cells Somatic cells of adults have 2 sets of chromosomes Haploid cells Gametes (egg and sperm)
Sexual Reproduction and Meiosis Chapter 11 1 Sexual life cycle Made up of meiosis and fertilization Diploid cells Somatic cells of adults have 2 sets of chromosomes Haploid cells Gametes (egg and sperm)
nutrients growth & division repellants movement
 Network Dynamics and Cell Physiology John J. Tyson Department of Biological Sciences & Virginia Bioinformatics Institute Outline 1. Cell Signaling: Physiology 2. Cell Signaling: Molecular Biology 3. Chemical
Network Dynamics and Cell Physiology John J. Tyson Department of Biological Sciences & Virginia Bioinformatics Institute Outline 1. Cell Signaling: Physiology 2. Cell Signaling: Molecular Biology 3. Chemical
Chapter 12: The Cell Cycle
 Name Period Chapter 12: The Cell Cycle Overview: 1. What are the three key roles of cell division? State each role, and give an example. Key Role Example 2. What is meant by the cell cycle? Concept 12.1
Name Period Chapter 12: The Cell Cycle Overview: 1. What are the three key roles of cell division? State each role, and give an example. Key Role Example 2. What is meant by the cell cycle? Concept 12.1
A. Incorrect! The Cell Cycle contains 4 distinct phases: (1) G 1, (2) S Phase, (3) G 2 and (4) M Phase.
 G 1, (2) S Phase, (3) G 2 and (4) M Phase.") Molecular Cell Biology - Problem Drill 21: Cell Cycle and Cell Death Question No. 1 of 10 1. Which of the following statements about the cell cycle is correct? Question #1 (A) The Cell Cycle contains 3
Molecular Cell Biology - Problem Drill 21: Cell Cycle and Cell Death Question No. 1 of 10 1. Which of the following statements about the cell cycle is correct? Question #1 (A) The Cell Cycle contains 3
3.2.2 All cells arise from other cells
 alevelbiology.co.uk SPECIFICATION Within multicellular organisms, not all cells retain the ability to divide. Eukaryotic cells that do retain the ability to divide show a cell cycle. DNA replication occurs
alevelbiology.co.uk SPECIFICATION Within multicellular organisms, not all cells retain the ability to divide. Eukaryotic cells that do retain the ability to divide show a cell cycle. DNA replication occurs
A diploid somatic cell from a rat has a total of 42 chromosomes (2n = 42). As in humans, sex chromosomes determine sex: XX in females and XY in males.
. As in humans, sex chromosomes determine sex: XX in females and XY in males.") Multiple Choice Use the following information for questions 1-3. A diploid somatic cell from a rat has a total of 42 chromosomes (2n = 42). As in humans, sex chromosomes determine sex: XX in females and
Multiple Choice Use the following information for questions 1-3. A diploid somatic cell from a rat has a total of 42 chromosomes (2n = 42). As in humans, sex chromosomes determine sex: XX in females and
Evolution of Ime2 Phosphorylation Sites on Cdk1 Substrates Provides a Mechanism to Limit the Effects of the Phosphatase Cdc14 in Meiosis
 Article Evolution of Ime2 Phosphorylation Sites on Cdk1 Substrates Provides a Mechanism to Limit the Effects of the Phosphatase Cdc14 in Meiosis Liam J. Holt, 1,2 Jessica E. Hutti, 3 Lewis C. Cantley,
Article Evolution of Ime2 Phosphorylation Sites on Cdk1 Substrates Provides a Mechanism to Limit the Effects of the Phosphatase Cdc14 in Meiosis Liam J. Holt, 1,2 Jessica E. Hutti, 3 Lewis C. Cantley,
DNA replication. M (mitosis)
") Introduction to cell cycle modeling Attila Csikász-Nagy G1 The cell cycle is the sequence of events whereby a growing cell replicates all its components and divides them more-or-less evenly between two
Introduction to cell cycle modeling Attila Csikász-Nagy G1 The cell cycle is the sequence of events whereby a growing cell replicates all its components and divides them more-or-less evenly between two
The Tem1 small GTPase controls actomyosin and septin dynamics during cytokinesis
 RESEARCH ARTICLE 1379 The Tem1 small GTPase controls actomyosin and septin dynamics during cytokinesis John Lippincott 1, *, Katie B. Shannon 1, *, Wenying Shou 2, Raymond J. Deshaies 2 and Rong Li 1,
RESEARCH ARTICLE 1379 The Tem1 small GTPase controls actomyosin and septin dynamics during cytokinesis John Lippincott 1, *, Katie B. Shannon 1, *, Wenying Shou 2, Raymond J. Deshaies 2 and Rong Li 1,
E. Incorrect! At telophase II, cells are nearly completed with meiosis, with no cross-over.
 OAT Biology - Problem Drill 06: Mitosis and Meiosis Question No. 1 of 10 1. During meiosis, cross-over between homologous chromosomes occurs at the end of. Question #01 (A) Anaphase II (B) Metaphase I
OAT Biology - Problem Drill 06: Mitosis and Meiosis Question No. 1 of 10 1. During meiosis, cross-over between homologous chromosomes occurs at the end of. Question #01 (A) Anaphase II (B) Metaphase I
Typical Life Cycle of Algae and Fungi. 5 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display.
 Module 3B Meiosis and Sexual Life Cycles In this module, we will examine a second type of cell division used by eukaryotic cells called meiosis. In addition, we will see how the 2 types of eukaryotic cell
Module 3B Meiosis and Sexual Life Cycles In this module, we will examine a second type of cell division used by eukaryotic cells called meiosis. In addition, we will see how the 2 types of eukaryotic cell
Chapter 6: Cell Growth and Reproduction Lesson 6.1: The Cell Cycle and Mitosis
 Chapter 6: Cell Growth and Reproduction Lesson 6.1: The Cell Cycle and Mitosis No matter the type of cell, all cells come from preexisting cells through the process of cell division. The cell may be the
Chapter 6: Cell Growth and Reproduction Lesson 6.1: The Cell Cycle and Mitosis No matter the type of cell, all cells come from preexisting cells through the process of cell division. The cell may be the
Cell Cycle Regulation by Chlamydomonas Cyclin-Dependent Protein Kinases
 Plant Cell Advance Publication. Published on February 5, 2018, doi:10.1105/tpc.18.00103 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 IN BRIEF Cell Cycle Regulation by Chlamydomonas
Plant Cell Advance Publication. Published on February 5, 2018, doi:10.1105/tpc.18.00103 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 IN BRIEF Cell Cycle Regulation by Chlamydomonas
Glc7/Protein Phosphatase 1 Regulatory Subunits Can Oppose the Ipl1/Aurora Protein Kinase by Redistributing Glc7
 MOLECULAR AND CELLULAR BIOLOGY, Apr. 2006, p. 2648 2660 Vol. 26, No. 7 0270-7306/06/$08.00 0 doi:10.1128/mcb.26.7.2648 2660.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved.
MOLECULAR AND CELLULAR BIOLOGY, Apr. 2006, p. 2648 2660 Vol. 26, No. 7 0270-7306/06/$08.00 0 doi:10.1128/mcb.26.7.2648 2660.2006 Copyright 2006, American Society for Microbiology. All Rights Reserved.
Cell Division (Outline)
") Cell Division (Outline) 1. Overview of purpose and roles. Comparison of prokaryotic and eukaryotic chromosomes and relation between organelles and cell division. 2. Eukaryotic cell reproduction: asexual
Cell Division (Outline) 1. Overview of purpose and roles. Comparison of prokaryotic and eukaryotic chromosomes and relation between organelles and cell division. 2. Eukaryotic cell reproduction: asexual
CELL CYCLE AND DIFFERENTIATION
 CELL CYCLE AND DIFFERENTIATION Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id WHAT IS CELL CYCLE? 09/12/14 d.purnomosari@ugm.ac.id
CELL CYCLE AND DIFFERENTIATION Dewajani Purnomosari Department of Histology and Cell Biology Faculty of Medicine Universitas Gadjah Mada d.purnomosari@ugm.ac.id WHAT IS CELL CYCLE? 09/12/14 d.purnomosari@ugm.ac.id
The Zds proteins control entry into mitosis and target protein phosphatase 2A to the Cdc25 phosphatase
 The Zds proteins control entry into mitosis and target protein phosphatase 2A to the Cdc25 phosphatase Sidonie Wicky John 1,2, Hendri Tjandra 1,3, David Schieltz 4, John Yates III 4, and Douglas R. Kellogg
The Zds proteins control entry into mitosis and target protein phosphatase 2A to the Cdc25 phosphatase Sidonie Wicky John 1,2, Hendri Tjandra 1,3, David Schieltz 4, John Yates III 4, and Douglas R. Kellogg
Chromosome duplication and distribution during cell division
 CELL DIVISION AND HEREDITY Student Packet SUMMARY IN EUKARYOTES, HERITABLE INFORMATION IS PASSED TO THE NEXT GENERATION VIA PROCESSES THAT INCLUDE THE CELL CYCLE, MITOSIS /MEIOSIS AND FERTILIZATION Mitosis
CELL DIVISION AND HEREDITY Student Packet SUMMARY IN EUKARYOTES, HERITABLE INFORMATION IS PASSED TO THE NEXT GENERATION VIA PROCESSES THAT INCLUDE THE CELL CYCLE, MITOSIS /MEIOSIS AND FERTILIZATION Mitosis
Review Quizzes Chapters 6-10
 Review Quizzes Chapters 6-10 1. The product(s) of the light reactions of photosynthesis is/are a. pyruvate b. glucose c. ATP and NADPH d. CO 2 and H 2 O e. ribulose bisphosphate (RuBP) 1. The product(s)
Review Quizzes Chapters 6-10 1. The product(s) of the light reactions of photosynthesis is/are a. pyruvate b. glucose c. ATP and NADPH d. CO 2 and H 2 O e. ribulose bisphosphate (RuBP) 1. The product(s)
cellular division cell division cell cycle cell cycle kinases chapter 18-19
 cellular division chapter 18-19 cell division when? growth replacement of older cells production of specialized cells asexual reproduction sexual reproduction production of gametes prokaryotic binary fission
cellular division chapter 18-19 cell division when? growth replacement of older cells production of specialized cells asexual reproduction sexual reproduction production of gametes prokaryotic binary fission
A phosphatase threshold sets the level of Cdk1 activity in early mitosis in budding yeast
 MBoC ARTICLE A phosphatase threshold sets the level of Cdk1 activity in early mitosis in budding yeast Stacy L. Harvey a, Germán Enciso b, *, Noah Dephoure c, Steven P. Gygi c, Jeremy Gunawardena b, and
MBoC ARTICLE A phosphatase threshold sets the level of Cdk1 activity in early mitosis in budding yeast Stacy L. Harvey a, Germán Enciso b, *, Noah Dephoure c, Steven P. Gygi c, Jeremy Gunawardena b, and
During meiosis, two consecutive nuclear divisions
 JCB: Article Cdc55 coordinates spindle assembly and chromosome disjunction during meiosis Farid Bizzari and Adele L. Marston Wellcome Trust Centre for Cell Biology, School of Biological Sciences, University
JCB: Article Cdc55 coordinates spindle assembly and chromosome disjunction during meiosis Farid Bizzari and Adele L. Marston Wellcome Trust Centre for Cell Biology, School of Biological Sciences, University
Cellular Division. copyright cmassengale
 Cellular Division 1 Cell Division All cells are derived from pre- existing cells New cells are produced for growth and to replace damaged or old cells Differs in prokaryotes (bacteria) and eukaryotes (protists,
Cellular Division 1 Cell Division All cells are derived from pre- existing cells New cells are produced for growth and to replace damaged or old cells Differs in prokaryotes (bacteria) and eukaryotes (protists,
Mitosis and Meiosis Cell growth and division
 Mitosis and Meiosis Cell growth and division The larger the cell, the more trouble the cell has moving nutrients and waste across the cell membrane. 1. DNA/information overload As a cell increases in size,
Mitosis and Meiosis Cell growth and division The larger the cell, the more trouble the cell has moving nutrients and waste across the cell membrane. 1. DNA/information overload As a cell increases in size,
Getting In and Out of Mitosis*
 Open Access NOBEL LAUREATE PERSPECTIVE Rambam Maimonides Medical Journal Getting In and Out of Mitosis* Tim Hunt, Ph.D., F.R.S. Nobel Laureate in Physiology or Medicine, 2001; Cancer Research UK, London
Open Access NOBEL LAUREATE PERSPECTIVE Rambam Maimonides Medical Journal Getting In and Out of Mitosis* Tim Hunt, Ph.D., F.R.S. Nobel Laureate in Physiology or Medicine, 2001; Cancer Research UK, London
Lecture #13 10/3 Dr. Wormington
 Lecture #13 10/3 Dr. Wormington The Molecular "Logic" of the Cell Cycle Recap 1. Cdks generally present throughout cell cycle but are inactive w/o cyclin subunits. 2. Cyclin subunits synthesized in discrete
Lecture #13 10/3 Dr. Wormington The Molecular "Logic" of the Cell Cycle Recap 1. Cdks generally present throughout cell cycle but are inactive w/o cyclin subunits. 2. Cyclin subunits synthesized in discrete
Bioinformatics 3. V18 Kinetic Motifs. Fri, Jan 8, 2016
 Bioinformatics 3 V18 Kinetic Motifs Fri, Jan 8, 2016 Modelling of Signalling Pathways Curr. Op. Cell Biol. 15 (2003) 221 1) How do the magnitudes of signal output and signal duration depend on the kinetic
Bioinformatics 3 V18 Kinetic Motifs Fri, Jan 8, 2016 Modelling of Signalling Pathways Curr. Op. Cell Biol. 15 (2003) 221 1) How do the magnitudes of signal output and signal duration depend on the kinetic
Cell Cycle Control in the Fission Yeast Schizosaccharomyces pombe
 Journal of Generul Microbiology ( 1989, 131, 2 123-2 127. Printed in Great Britain 2123 Cell Cycle Control in the Fission Yeast Schizosaccharomyces pombe The Tenth Fleming Lecture ByPAUL NURSE Imperial
Journal of Generul Microbiology ( 1989, 131, 2 123-2 127. Printed in Great Britain 2123 Cell Cycle Control in the Fission Yeast Schizosaccharomyces pombe The Tenth Fleming Lecture ByPAUL NURSE Imperial
3 RFC1-like clamps (Ctf18, Elg1 and Rad24) have nonessential and overlapping functions as seen by the increased sensitivity to DNA damage or slowed
 have nonessential and overlapping functions as seen by the increased sensitivity to DNA damage or slowed") 3 RFC1-like clamps (Ctf18, Elg1 and Rad24) have nonessential and overlapping functions as seen by the increased sensitivity to DNA damage or slowed replication Initiation of Replication http://www.orst.edu/instruction/bb492/figletters/figh3.gif
3 RFC1-like clamps (Ctf18, Elg1 and Rad24) have nonessential and overlapping functions as seen by the increased sensitivity to DNA damage or slowed replication Initiation of Replication http://www.orst.edu/instruction/bb492/figletters/figh3.gif
Bioinformatics 3! V20 Kinetic Motifs" Mon, Jan 13, 2014"
 Bioinformatics 3! V20 Kinetic Motifs" Mon, Jan 13, 2014" Modelling of Signalling Pathways" Curr. Op. Cell Biol. 15 (2003) 221" 1) How do the magnitudes of signal output and signal duration depend on the
Bioinformatics 3! V20 Kinetic Motifs" Mon, Jan 13, 2014" Modelling of Signalling Pathways" Curr. Op. Cell Biol. 15 (2003) 221" 1) How do the magnitudes of signal output and signal duration depend on the
Roles of Cell Division. Reproduction - Like begets like, more or less. Examples of Cell Numbers. Outline Cell Reproduction
 Outline Cell Reproduction 1. Overview of Cell Reproduction 2. Cell Reproduction in Prokaryotes 3. Cell Reproduction in Eukaryotes 1. Chromosomes 2. Cell Cycle 3. Mitosis and Cytokinesis 4. Sexual Life
Outline Cell Reproduction 1. Overview of Cell Reproduction 2. Cell Reproduction in Prokaryotes 3. Cell Reproduction in Eukaryotes 1. Chromosomes 2. Cell Cycle 3. Mitosis and Cytokinesis 4. Sexual Life
Budding Yeast Greatwall and Endosulfines Control Activity and Spatial Regulation of PP2A Cdc55 for Timely Mitotic Progression
 Budding Yeast Greatwall and Endosulfines Control Activity and Spatial Regulation of PP2A Cdc55 for Timely Mitotic Progression Maria Angeles Juanes 1., Rita Khoueiry 1., Thomas Kupka 2, Anna Castro 1, Ingrid
Budding Yeast Greatwall and Endosulfines Control Activity and Spatial Regulation of PP2A Cdc55 for Timely Mitotic Progression Maria Angeles Juanes 1., Rita Khoueiry 1., Thomas Kupka 2, Anna Castro 1, Ingrid
2005 LANDES BIOSCIENCE. DO NOT DISTRIBUTE.
 [Cell Cycle 4:8, 1093-1098; ugust 2005]; 2005 Landes ioscience Report FER but not MEN Genes are Required for Exit from Meiosis I Rebecca J. Kamieniecki 1,2 Li Liu 1 Dean S. Dawson 1,2, * 1 Department of
[Cell Cycle 4:8, 1093-1098; ugust 2005]; 2005 Landes ioscience Report FER but not MEN Genes are Required for Exit from Meiosis I Rebecca J. Kamieniecki 1,2 Li Liu 1 Dean S. Dawson 1,2, * 1 Department of
Chromosome Chr Duplica Duplic t a ion Pixley
 Chromosome Duplication Pixley Figure 4-6 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-72 Molecular Biology of the Cell ( Garland Science 2008) Interphase During mitosis (cell division),
Chromosome Duplication Pixley Figure 4-6 Molecular Biology of the Cell ( Garland Science 2008) Figure 4-72 Molecular Biology of the Cell ( Garland Science 2008) Interphase During mitosis (cell division),