A Leucine Zipper Motif Essential for Gating of Hyperpolarization-Activated Channels*
|
|
|
- Joella Sims
- 6 years ago
- Views:
Transcription
1 JBC Papers in Press. Published on October 9, 2012 as Manuscript M The latest version is at A leucine zipper essential for HCN gating A Leucine Zipper Motif Essential for Gating of Hyperpolarization-Activated Channels* Konstantin Wemhöner 1,#, Nicole Silbernagel 1,#, Stefanie Marzian 1, Michael F. Netter 1, Susanne Rinné 1, Phillip J. Stansfeld 2, and Niels Decher 1,3 1 From the Institute for Physiology, Vegetative Physiology Group, University of Marburg, Deutschhausstrasse 1-2, Marburg, Germany 2 Structural Bioinformatics and Computational Biochemistry Unit, Department of Biochemistry, University of Oxford, South Parks Road, Oxford OX1 3QU, UK *Running Head: A leucine zipper essential for HCN gating 3 To whom correspondence should be addressed: Niels Decher, PhD, Philipps University Marburg, Institute for Physiology and Pathophysiology, Vegetative Physiology, Deutschhausstrasse 1-2, D Marburg, Germany; Fax: (+49) ; decher@staff.uni-marburg.de # These authors contributed equally to this work Keywords: Leucine zipper: hyperpolarization-activated: HCN2 Background: It is poorly understood how hyperpolarization-activated cyclic nucleotidegated channels (HCNs) function. Results: We have identified a leucine zipper in the S5 segment of HCNs, regulating hyperpolarization-activated and instantaneous current components. Conclusion: The leucine zipper is essential for HCN channel gating. Significance: The identification and functional characterization of the leucine zipper is an important step towards the understanding of HCN channel function. SUMMARY Hyperpolarization-activated cyclic nucleotide-gated (HCN) channels are pacemakers in cardiac myocytes and neurons. Although their membrane topology closely resembles that of voltage-gated K + channels, the mechanism of their unique gating behavior in response to hyperpolarization is still poorly understood. We have identified a highly conserved leucine zipper motif in the S5 segment of HCN family members. In order to study the role of this motif for channel function, the leucine residues of the zipper were individually mutated to alanine, arginine or glutamine residues. Leucine zipper mutants traffic to the plasma membrane, but the channels lose their sensitivity to open upon hyperpolarization. Thus, our data indicate that the leucine zipper is an important molecular determinant for hyperpolarization-activated channel gating. Residues of the leucine zipper interact with the adjacent S6 segment of the channel. This interaction is essential for voltage-dependent gating of the channel. The lower part of the leucine zipper, at the intracellular mouth of the channel, is important for stabilizing the closed state. Mutations at these sites increase current amplitudes or result in channels with deficient closing and increased min-po. Our data are further supported by homology models of the open and closed state of the HCN2 channel pore. Thus, we conclude that the leucine zipper of HCN channels is a major determinant for hyperpolarization-activated channel gating. Following an action potential, a slow spontaneous depolarization drives specialized pacemaking cells in cardiac and neuronal tissues towards another action potential. This pacemaker current, called I f in cardiac cells and I h in the nervous system, is mainly carried by channels referred to as hyperpolarization-activated cyclic nucleotide-gated (HCN) channels (1-2). The HCN channel family comprises four members (3-4). The membrane topology of HCN channels closely resembles that of voltage-gated K + (Kv) channels, as they also have six transmembrane segments (S1-S6), a pore loop with a selectivity filter and a charged S4 segment acting as a voltage-sensor. In contrast to Kv channels, HCN channels open in response to hyperpolarization and are also permeable to Na + (2). The mechanism for this uncommon gating has been the subject of many studies (5-9). However, the 1 Copyright 2012 by The American Society for Biochemistry and Molecular Biology, Inc.
2 inverse gating phenotype of HCN channels is still poorly understood. Leucine zipper motifs are found in alphahelical parts of proteins, mediating the interaction between two alpha-helical domains. Leucine zippers were initially described as highly conserved motifs involved in the interaction of transcription factors with DNA (10). As the interaction of two opposed leucine zippers leads to stabilized, more rigid proteins, the zippers can be of relevance for any form of protein-protein assembly. The motif is x-(leu-x-x-y-x-x-x-) n - Leu, where x can be any amino acid, Y is mostly an aliphatic residue and n is the number of repeats, with at least three. While the aforementioned motif is the classical leucine zipper, there is also a modified leucine zipper motif, where any leucine can be replaced by isoleucine or valine. Leucine zippers in ion channels were previously identified to be of major relevance for several different functions. A leucine zipper in the S4-S5 linker of the drosophila Shaker channel is important for the coupling of S4 movement to opening of the inner pore helix (11). The Ca 2+ activated hik1 channels have a C-terminal leucine zipper essential for proper folding and trafficking of the channel (12). Calcium channels of the Ca V 1.x-family assemble with the A-kinaseanchoring protein AKAP15 depending on a leucine zipper in the distal C-terminus (13-14). The binding of AKAP15 scavenges protein kinase A to the channel, resulting in increased calcium current amplitudes (14). In CNG channels, which are closely related to HCN channels, a leucine zipper following the cyclic nucleotide binding domain (CNBD) is required for regulating the stoichiometry of CNG channel assembly (15). Using a bioinformatic approach, we have identified a leucine zipper motif within the S5 segment of HCN channels. The leucine zipper is highly conserved and is present in all HCN channel family members and in sphcn channels. We used site-directed mutagenesis, electrophysiological recordings, surface expression analysis and molecular modeling to determine the structural and functional role of the leucine zipper motif in HCN2. Our data indicate that the leucine zipper is essential for an interaction of the S5 and S6 segments and that this interaction is important for proper gating of HCN channels. EXPERIMENTAL PROCEDURES Molecular biology We used a mouse HCN2 clone with a deletion of the GC-rich N-terminus (amino acids 2-130), which we have previously described as nthcn2 (5). HCN1 was studied using mouse HCN1 cloned in the pbf1 vector. QuikChange (Stratagene) site directed mutagenesis kit was used to introduce mutations into the nthcn2 cdna or mhcn1 cdna. Complementary RNA (crna) for injection into Xenopus oocytes was prepared with mmessage mmachine SP6 Kit (Ambion) after linearization with EcoRI (HCN2) or MluI (HCN1) (both Fermentas). To assess the surface expression of HCN channels, a modified hemagglutinin (HA) protein epitope was inserted in HCN2 as previously described (5). Expression of HCN channels in oocytes Stage IV and V oocytes from mature Xenopus laevis toads were isolated and injected with 3.5 ng of crna of either wild-type or mutant HCN2 channels for electrophysiological recordings or with 10 ng of crna for surface expression assay. For HCN1 7.5 ng of wild-type or mutant crna was injected. Isolated oocytes were stored at 18 C in ND96 recording solution (containing in mm: NaCl 96, KCl 2, CaCl 2 1.8, MgCl 2 1, HEPES 5, ph 7.4) with added Na-pyruvate (275 mg/l), theophylline (90 mg/l) and gentamicin (50 mg/l). Two-electrode voltage clamp recordings Standard two-electrode voltage-clamp experiments (TEVC) were performed at room temperature (21 22 C) in ND66 recording solution (containing in mm: NaCl 66, KCl 32, CaCl 2 1.8, MgCl 2 1, HEPES 5, ph 7.4) 2 to 3 days after injection of oocytes with crna. Microelectrodes were fabricated from glass pipettes filled with 3 M KCl and had a resistance of MΩ. Currents were elicited by 2 s pulses applied in 10 mv decrements to potentials ranging from +30 mv to -140 mv followed by a 750 ms test pulse to -130 mv. Normalized tail current amplitude taken from the 750 ms test pulse to -130 mv was plotted versus the test potential to obtain the relative conductancevoltage (G-V) relationship and fitted with a Boltzmann function: I t = (1 min-p o )/[1 + exp((v - V 1/2 )/k)] + min-p o where V 1/2 is the voltage required for halfmaximal activation, k is the slope factor, a measure of the voltage dependence of channel gating and min-p o is the minimum open probability defined as the fraction of channels which are instantaneously open. Surface expression assay in Xenopus oocytes A chemiluminescence assay was performed as described by Zerangue et al. (16). 2
3 Briefly, oocytes were blocked with ND96/1% bovine serum albumin, labeled with anti-ha antibody, sequentially washed with ND96 solution (containing in mm: NaCl 96, KCl 2, CaCl 2 1.8, MgCl 2 1, HEPES 5; ph 7.4 with NaOH). For detection horseradish peroxidaseconjugated secondary antibody was used. Individual oocytes were placed in 20 μl SuperSignal Elisa Femto solution (Thermo Scientific, Rockford, IL, USA) and, after an equilibration period of 10 s, chemiluminescence was quantified in a luminometer (GloMax 20/20, Promega, Madison, WI, USA). The surface expression of oocytes per construct (from three different batches of oocytes) was analyzed. Non-injected oocytes served as negative control. The difference between injected and non-injected oocytes was determined and the relative change in the luminescence signal was compared for the different HA-tagged HCN2 mutant constructs. Western blot analysis Twenty oocytes per construct were homogenized in 400 µl lysis buffer (containing in mm: NaCl 150, Tris/HCl 20, ph 7.5, 1% Triton-X-100). Insoluble material was separated by centrifugation for 15 min at 4 C. The protein content of the supernatants was quantified and subsequently probes were heatdenatured in sample buffer at 50 C for 10 min. Similar amounts of total protein were separated for each probe on 12% SDS polyacrylamide gels and visualized by immunoblotting with rat monoclonal anti-ha primary antibodies (clone 3F10, 1:1000 dilution; Roche Pharmaceuticals). The binding of the primary antibodies was detected using peroxidase-conjugated goat antirat antibodies (Jackson ImmunoResearch) and a chemiluminescent extended-duration substrate, Super Signal West Dura (Thermo Scientific). Homology modeling Modeller 9v9 (17) was used to create a closed homology model of the pore domain and S4-S5 linker of mouse HCN2 from the MlotiK1 crystal structure (18) [PDB ID: 3BEH]. The similarity between the two sequences was 29.2%. An open state model was also built, with the inner S6 helices modelled using the open state Kv1.2:Kv2.1 chimera (KvChim) [PDB ID: 2R9R] crystal structure (19) as a template. Four-fold symmetry was applied to both models. The final HCN2 models were compared to their crystal structure template to verify that the modeling step had not significantly altered backbone and side chain conformations. Data analyses Data are reported as means ± S.E.M. Statistical significance was calculated using unpaired Student s t-test. All experiments 3 were performed at room temperature (22-24 C). *, indicates P < 5; **, P < 1; P < 01. n.s. indicates non-significant changes. RESULTS A conserved leucine zipper motif in hyperpolarization-activated channels We have identified a leucine zipper motif in the S5 segment of HCN channels (Fig. 1, A and B). The leucine zipper is conserved in hyperpolarizationactivated cation channels from different species, including sphcn channels from sea urchin (Fig. 1A). Parts of the leucine zipper motif are also present in the closely related CNG channels (Fig. 1A). We have functionally analyzed the leucine zipper in mouse HCN2 channels, with zipper residues located at L343, L350, L357 and L364 (Fig. 1, A and B). Alanine mutations at the leucine zipper motif of HCN2 lead to decreased current amplitudes due to altered channel gating For the current study we used an HCN2 channel construct with an N-terminal truncation which was previously used in functional studies of HCN gating (5-6,9). Currents elicited by HCN2 expressed in Xenopus oocytes showed typical characteristics (Fig. 2A). We have analyzed the current amplitudes of channels with leucine zipper residues mutated to alanine. Therefore, we injected 3.5 ng crna of wild-type HCN2, L343A, L350A, L357A and L364A. The current amplitudes were measured 48 hours after injection. Currents of wild-type HCN2 channels were 7.2 µa ± 0.4 µa (n = 46; normalized 0 ± 5). Although the leucine zipper alanine mutants generated hyperpolarization-activated currents, the amplitude was strongly reduced for all zipper mutant constructs (Fig. 2, A and B and supplemental Fig. S1, A and B). The higher the location of the leucine zipper mutation in the S5 segment, the more pronounced was the reduction in current amplitudes. Current size was HCN2 > L343A > L350A > L357A > L364A (Fig. 2B and supplemental Fig. S1B). The observed reduction of current amplitudes may be due to a reduced protein expression, a deficient channel trafficking or an altered channel gating. To further clarify the mechanism of current reduction, we performed western blot experiments and a chemiluminescence based surface expression assay. For these experiments an HA-tag was introduced in the extracellular S3-S4 loop between Gly-284 and Ile-285. Western blot analysis of oocytes injected with HCN2-HA constructs revealed that similar amount of channel protein was present for
4 all the zipper mutants (Fig. 2C). However, while current amplitudes are reduced, the chemiluminescence-assay revealed that the surface expression of mutant channels was similar as for wild-type channels (Fig. 2D). Thus, current reduction of the zipper mutants does not result from altered channel trafficking and must be caused by an altered conductivity or voltage dependence of HCN2 channels. Changes in voltage dependence of activation by leucine zipper alanine mutations The voltage dependence of channel activation was determined by analyzing the instantaneous tail current at -130 mv. The V 1/2 of activation for wild-type HCN2 was ± 0.3 mv and slope factor was 8.7 ± 0.25 (n = 10). G/V-curves for L343A, L350A and L357A were similar as for wild-type channels (Fig. 2E, Table 1 and supplemental Fig. S1C). All constructs (except L364A) had only minor shifts of the V 1/2 of activation (Fig. 2E, Table 1 and supplemental Fig. S1C). Due to a steeper slope factor k (Table 1), L350A reached its maximum current at around -100 mv, while the other mutants (except L364A) reached steadystate activation at about -120 mv (Fig. 2E). Slope factors were significantly changed for L350A (5.6 ± 0.2) and L364A (17.2 ± 1.3), indicating an altered voltage dependence of channel gating for these constructs (Table 1). In contrast to other leucine zipper alanine mutations L364A showed only a little hyperpolarization-activated current component, with a V 1/2 of activation that is more negative than -140 mv (Fig. 2E, Table 1 and supplemental Fig. S1C). Summarizing, L343A, L350A and L357A have reduced current amplitudes with only minor changes in the voltage dependence of activation, while L364A currents are strongly reduced due to an uncoupling, as the channels open only upon very strong hyperpolarization of the membrane. Double and triple alanine mutations lead to a strong reduction in current amplitudes and negative shifts in voltage dependence of activation Next, we mutated multiple leucine residues at once and generated the doublemutations L343A/L350A, L343A/L357A, L350A/L357A and the triple mutant L343A/L350A/L357A. The uppermost S5 leucine zipper mutation L364A was not included in this multiple mutation approach as its currentvoltage relationship was already shifted to very negative potentials. All two- and three-fold mutants showed functional hyperpolarizationactivated HCN currents when expressed in Xenopus oocytes (Fig. 3A), but current amplitudes were strongly reduced (Fig. 3B). 4 Again, current reductions were more pronounced for mutations in the upper part of the S5 segment. Current size was HCN2 > L343A/L350A > L343A/L357A > L350A/L357A (Fig. 3B). Our chemiluminescence assays revealed that the double mutants L343A/L350A and L343A/L357A, efficiently traffic to the surface membrane (Fig. 3C), while TEVC recordings reveal that they have an altered channel gating, as they fail to open upon hyperpolarization (Fig. 3D). In contrast, the L350A/L357A double and the L343A/L350A/L357A triple mutants have a strongly reduced surface expression (Fig. 3C). Note that similar efficiency in protein expression was detected by western blot analysis for all constructs (Fig. 3C, inset). These data further highlight the relevance of the zipper residues in HCN channel gating, but also indicate that the leucine zipper might be important for the formation of a stable HCN channel complex, necessary for the subsequent trafficking to the surface membrane. Of the single alanine mutants, only the uppermost S5 segment residue L364A showed a deficiency in opening upon hyperpolarization (Fig. 2E and supplemental Fig. S1C). While the single mutations L343A, L350A and L357A had only small changes in voltage dependence (Fig. 2E and supplemental Fig. S1C), the double mutations L343A/L357A and L350A/L357A showed drastic shifts in the voltage dependence of activation (Fig. 3D and Table 1). For the lower-most S5 double mutation, L343A/L350A, we observed only a minor change in the slope of the voltage dependence (Fig. 3D and Table 1), but a significantly higher min-po (0.177 ± 13, n = 8) (Fig. 3D), indicating more instantaneous currents, as a measure of a closing deficiency. While this L434A/L350A mutant had no strong shift in the voltage dependence of activation, introducing a third alanine mutation (L357A) into this construct (L343A/L350A/L357A) resulted in channels that open only upon strong hyperpolarization of the plasma membrane (Fig. 3D). Summarizing, current reduction and shift of the voltage dependence of activation are more pronounced for residues of the upper part of the leucine zipper. Multiple alanine mutations lead to an impaired voltage dependence of HCN channel gating and for the lower part of the leucine zipper we observed changes in the closing of the channel (increased min-po of L343A/L350A). Mutations of leucine zipper residues to glutamine or arginine strongly reduce current amplitudes and voltage dependence of HCN
5 channels As a leucine to alanine mutation is a conservative mutation, we studied the effects of mutating leucine zipper residues to a bulky and charged (arginine) or to a bulky and aliphatic (glutamine) residue. Representative current traces of arginine and glutamine mutants of the leucine zipper are illustrated in Figure 4A and 4F. Arginine and glutamine mutations at residues L350, L357 and L364 led to an even more pronounced reduction in current amplitudes (Fig. 4, A-B and F-G) when compared to the respective alanine mutants (Fig. 2, A and B). In contrast to the current reduction by the L343A mutation (Fig. 2, A and B), a mutation to a bulky amino acid resulted in increased current amplitudes. Mutation of the leucine residue at position 343 led to 2.1 ± 0.2 fold (L343R) and 6.1 ± 0.6 fold (L343Q) increased current amplitudes (Fig. 4, B and G). The western blot and surface expression experiments revealed that all mutants are translated with the same efficiency (Fig. 4, C and H) and have a similar trafficking to the plasma membrane (Fig. 4, D and I). Thus, as for the alanine mutants, the effects on current amplitudes cannot be explained by changes in protein expression of the mutants or by an altered trafficking. Next, we analyzed the voltage dependence of the arginine and glutamine mutants. L350Q and L350R were not studied, as these did not open at all upon hyperpolarization (Fig. 4, A and F). Arginine and glutamine mutations at L357 and L364 shifted the voltage of half-maximal activation to very hyperpolarized potentials (Fig. 4, E and J and Table 1). Consistent with the single and double alanine mutants, we did not observe a strong negative shift in the voltage dependence for mutants at the lowest S5 segment residue L343. We found that the current voltage relationship of L343Q channels did not differ from that of wild-type HCN2 (Fig. 4E). For L343R the V 1/2 was even shifted by about +17 mv to more positive potentials (Fig. 4J and Table 1). Interestingly, we observed a massive increase in min-po for L343R (Fig. 4, F and J). Note that a significant rise in min-po was also observed for the lowest S5 zipper double mutant (L343A/L350A) (Fig. 3D). These results indicate that all residues of the leucine zipper are important for proper channel function upon hyperpolarization with the L343 residue having a special role in regulating the open state of the channel. Experiments with HA-tagged and full-length HCN2 constructs confirm the functional role of the leucine zipper in HCN2 gating Next, we 5 performed control experiments to verify that the extracellular HA-tag introduced in HCN2 does not interfere with the gating phenotype of the mutants or the trafficking of the channels. Note that the HA-tagged wild-type HCN2 is activating at more hyperpolarized potentials and is slower in activation as the non-tagged HCN2 (supplemental Fig. S2A). However, the HAtagged HCN2 constructs, used in Fig. 4, H and I revealed the same electrophysiological phenotypes (supplemental Fig. S2, A-C) as the untagged HCN2 channel constructs (Fig. 4, F-G and J). The L343R mutant again shows a rightward shift in the voltage dependence of activation (supplemental Fig. S2C), an increased min-po (supplemental Fig. S2, A and C) and a gain-of-function in the current amplitude (supplemental Fig. S2, A and B). In accordance to the untagged channels, the mutants L350R, L357R and L364R also show reduced current amplitudes in the HA-background and shifts of the voltage-dependence out of the physiological range (supplemental Fig. S2, A-C). As the relative current amplitudes also match with the untagged versions of the mutants, the HA-epitope most likely does not interfere with the trafficking of the channel. To rule out that the N-terminal deletion of the nthcn2 construct (5-6,9) used in the current study interferes with the channel gating of the zipper mutants, we studied the arginine mutants described in Figure 4 in the full-length HCN2 channel. The data obtained for the full-length HCN2 were very similar to that using the nthcn2 (supplemental Fig. S2D). Thus, neither the N-terminal deletion nor the HA-epitope interfered with the results obtained with the nthcn2 channel construct. The S5 segment leucine zipper interacts with an opposing S6 segment If the leucines in the S5 segment of HCN channels act as a zipper motif, these residues should interact with aliphatic side chains of an opposing helix to form a stable structure. To identify a domain and residues that interact with the leucine zipper in the S5 segment, we developed a closed state homology model of the mouse HCN2 channel based on the crystal structure of the closely related CNG channel MlotiK1 (18). The pore homology model included the S4-S5 linker, the S5 and S6 segments (Fig. 5A). The model predicted an interaction of the leucine zipper residues with opposing aliphatic residues of the S6 segment from the same subunit (Fig. 5A). L343 was predicted to interact with L438, L350 with M430, L357 with V423 and L364 was predicted to
6 interact with L416. Therefore, we hypothesized that the leucine zipper is essential for the functional packing of channel proteins and that it might play a role in the conversion from the closed to the open state of HCN channels. To test for a putative interaction of the leucine residues with the residues in the S6 segment, we analyzed alanine mutations at the residues L416, V423, M430 and L438A (Fig. 5B). The current size was reduced in a similar manner as for the S5 segment alanine mutants. Meaning, the reduction in current amplitude was more pronounced for alanine mutations located in the upper S6 segment, similar as observed for the S5 segment (Fig. 5C). The current reduction by the S6 segment mutations L416A and L438A was also confirmed in the full-length HCN2 channel (supplemental Fig. S3). L438 which is located in the lower S6 segment, is supposed to interact with L343. Interestingly, L438A also shows an increased instantaneous current component (Fig. 5B and arrow in supplemental Fig. S3A), similar as L343R or L343A/L350A. Taken together, these results support a possible interaction of the S5 and S6 domains via the leucine zipper and the residues proposed by our model. Mutations in HCN1 confirm the relevance of leucine zippers for the HCN channel family As the leucine zipper motif is highly conserved within the HCN channel family (Fig. 1A), we studied glutamine mutations in the leucine zipper of HCN1. Current amplitudes of the L290Q, L297Q, L304Q and L311Q HCN1 mutants were strongly reduced (Fig. 6, A and B and supplemental Fig. S4), similar as for the HCN2 glutamine mutants (Fig. 4, A and B). HCN1 leucine zipper mutants only open upon very strong hyperpolarization of the plasma membrane (Table 2). However, while the lowest S5 zipper mutation in HCN2 L343Q resulted in a gain-offunction (Fig. 4B), the lowest HCN1 zipper mutation L290Q resulted in a current reduction (Fig. 6, A and B). Thus, we analyzed the current amplitudes of different residues introduced at position L343 of HCN2. As the increased current amplitudes were specific for arginine and glutamine residues at this position (Fig. 6C), we conclude that the gain-of-function needs a large neutral or positively charged residue at position L343 of HCN2. In contrast, HCN1 currents were strongly reduced by arginine and glutamine residues introduced at the homologous site (L290). Thus, similar as for the leucine zipper in HCN2, the lowest zipper residue of HCN1 seems to be involved in regulating the open and closing 6 properties of the channel, albeit in a different manner (see Discussion section). Note that the lowest HCN1 zipper mutant, L290Q, has an increased min-po (Fig. 6A, arrow). Similar effects were observed for the L343A and L343R mutations in HCN2 (Figs. 2A and 4F). The data obtained using the HCN1 channel further highlight the functional relevance of the leucine zipper for HCNs to function as hyperpolarization-activated channels and indicate that the lowest S5 zipper residue is involved in regulation the open state of the channel. The lowermost leucine zipper residue in the S5 segment regulates the open state of HCNs As we have observed increased instantaneous current amplitudes (increased min-po) for mutations at the lowermost S5 leucine zipper residue, we studied the instantaneous current of channels mutated at this and the corresponding S6 site in more detail. Wild-type HCN1 currents have about 20 % of instantaneous currents when activated by a voltage step to -110 mv (Fig. 7A, grey trace and arrow). In contrast, for L290Q about 60 % of the current was already instantaneously activated (Fig. 7A, grey trace and arrow). For L290R 100 % of the channels were already in the open state when membrane potential was stepped to -110 mv (Fig. 7A). Wild-type HCN2 currents show about 8 % of instantaneous currents when activated by a voltage step to -110 mv (Fig. 7B). Similar to the HCN1 mutants, HCN2 channel mutants at the lowermost S5 zipper residue generate currents with an increased instantaneous component (Fig. 7B). The instantaneous current component was 35 % for L343A and 60 % for L343R (Fig. 7B). Introducing similar mutations at the corresponding S6 site L438, also results in increased instantaneous current components (Fig. 7B). The instantaneous current was 60 % for L438A and 70 % for L438R (Fig. 7B). These data support our hypothesis that the first residue of the leucine zipper, the lowermost in the S5 segment, regulates the open state of HCN channels. In addition, it further supports an interaction of L343 of the S5 with L438 of the S6 segment. This interaction might stabilize the closed state of the channel as suggested by our closed state pore homology model (Fig. 5A). Instantaneously open mutants of HCN2 and HCN1 have unaltered ion selectivity As mutations at the lowermost leucine zipper residue and the pairing S6 segment site lead to currents with leak channel characteristics, we analyzed whether these mutations alter the ion selectivity of HCN2 and HCN1. Therefore, we recorded
7 fully-activated current-voltage relationships of HCN2 (L438R) and HCN1 (L290R) (Fig. 8, A and B). For both constructs the reversal potentials were similar to that of the respective wild-type channels, indicating that the ion selectivity was not changed for both channel isoforms. Thus, neither the S5 mutant (HCN1-L290R) nor the S6 mutant (HCN2-L438R) caused a major change in ion selectivity. For the L290R mutant however, we have observed a change in the rectification pattern of the fully-activated current voltage relationship (Fig. 8B), further supporting our observation that mutations at the lowest leucine zipper residue can induce an altered channel gating. In addition, it was apparent from the fully-activated current-voltage relationship recordings that both mutants, L438R and L290R, have a slower rate of deactivation, indicating that the mutations interfere with proper closing of the channels. The lowermost part of the leucine zipper stabilizes a salt-bridge and the closed state of HCN2 To analyze the role of an interaction of the lower leucine zipper with the late S6 segment during channel gating, we generated an open pore homology model of HCN2 based on the rkv1.2 crystal structure (19), for details see Methods section. In the closed state model the S5 residue L343 is in close proximity to L438 of the S6 segment (Fig. 9, A and C). In contrast, due to the movement of the S6 segment during channel opening, L438 dislodges from L343 (Fig. 9, B and D). Thus, an interaction of these residues might stabilize only the closed state of the channel. We have previously described that a salt-bridge between the S4-S5 linker residue R339 and the late S6 segment residue D443 stabilizes the closed state of the channel (9). Our closed state and open state homology models are in good agreement with these findings, as the salt-bridge is favored in the closed state model (Fig. 9, C and D). Opening of the channel leads to a sideward movement and rotation of the S6 segment, displacing D443 from R339 (Fig. 9, C and D). From our modeling data, we again conclude that the open state of HCN2 is regulated by a salt-bridge of the S4-S5 linker and the late S6 segment. We propose that this salt bridge is stabilized by an interaction of the lower leucine zipper (L343) interacting with the S6 segment (L438). Mutations at the zipper residue L343 or the S6 residue L438 might de-stabilize the closed state. In addition, bulky amino acids introduced at position L438 (Fig. 9D) might sterically hinder the downward movement of the S6 segment and the returning to the closed state of the channel. DISCUSSION It is still not fully understood how voltage sensor movement in hyperpolarization-activated channels is coupled to channel opening and closing. For Kv channels a leucine zipper motif in the S4-S5 linker was described as an essential determinant for the transduction of charge movement into channel gating (11). Thus, when we observed a highly conserved leucine zipper motif in the S5 segment it was prompting to analyze its role for HCN channel function. Our data indicate that the leucine zipper is an important molecular determinant for hyperpolarization-activated channel gating. Our modeling data predicted an interaction of the leucine zipper in the S5 segment with the adjacent S6 segment of HCN channels. This interaction appears to be essential for voltagedependent gating of hyperpolarization-activated channels. We found that alanine, glutamine and arginine mutations of the leucine zipper residues lead to a strong reduction in HCN current amplitudes (with the exception of L343Q/R). The higher the leucine zipper mutations were located in the S5 segment the more pronounced was the reduction in current amplitudes. When single or double alanine mutations were located high in the S5 segment, the channels were no longer able to open upon hyperpolarization. All arginine and glutamine mutations of the leucine zipper generated channels which did not open upon hyperpolarization, except for mutations in the lower part of the S5 segment. The lower part of the leucine zipper, at the intracellular mouth of the channel, appears to be important for the stabilization of the closed state of the channel. Arginine and glutamine mutations at these sites increased current amplitudes or resulted in channels with deficient closing and an increased instantaneous current component (min-po). Summarizing, the upper part of the leucine zipper might be important for stabilizing a rigid S5 and S6 segment interaction which is necessary for a proper hyperpolarization-activated channel gating at the intracellular channel mouth. The lower part of the leucine zipper might be more flexible, as it is directly located at the inner channel gate formed by the S4-S5 linker, the early S5 and late S6 segment. Here leucine zipper residues might influence the open and closed state of the channels. 7
8 It is known that potassium channel gating needs global rearrangements of the S4, S5 and S6 domains. Voltage-dependent gating can be introduced into KcsA channels by constructing KcsA-Shaker chimeras (20). Here, voltagedependent gating is critically dependent on the interaction of parts of the S4-S5 linker with residues in the late S6 segment (20). However, also concrete interactions or salt bridges between domains seem to be of major relevance for channel gating (8-9). In HERG channels which are more closely related to HCN than to Kv channels, a single charge-reversing mutation in the S4-S5 linker (D540K) is sufficient to prevent channel closure (21). In addition, the D540K mutant channels are able to open upon depolarization or hyperpolarization. The opening response to hyperpolarization can be abolished by introducing a second mutation at position R665 in the late S6 segment of HERG (8). The authors suggest a salt-bridge between D540 and R665 and that repulsion between K540 and R665 mediates the HCN-like channel gating in D540K HERG. A corresponding salt-bridge of the S4-S5 linker with the late S6 was proposed for HCN channels (9). An arginine residue in the S4-S5 linker of HCN2 (R339) was found to interact with an aspartate in the late S6 or early C-linker (D443). This interaction stabilizes the closed state of wild-type channels by an electrostatic interaction. Mutating R339 in the S4-S5 linker and D443 in the early C-linker de-stabilizes the closed state of the channel, resulting in instantaneous HCN currents. We have observed a similar phenotype for arginine and glutamine mutations at the lowermost S5 leucine zipper residue L343 and the S6 residue L438 which is predicted to interact with L343. We conclude that the lower part of the leucine zipper stabilizes the closed state of the channel. According to our state dependent HCN models, the residue R339 (S4-S5 linker) forms a salt-bridge with D443 (late S6 or C-linker) to stabilize the closed state. Thus, our models support the previously proposed electrostatic interaction (9). Most importantly, this salt-bridge might be stabilized by the lowest leucine zipper residue L343, which is preferentially interacting with the S6 residue L438 when the channels are in the closed state. Accordingly, mutations of either of the residues (R339 and D443 or L343 and L438) result in a de-stabilization of the closed state and in instantaneous currents. Our findings support the hypothesis of a concrete interaction of amino acids at the intracellular mouth to modulate HCN channel gating. Interactions of residues in the 8 S4-S5 linker with the C-linker and of the early S5 with the late S6 stabilize the closed state of the channel. Despite the lack of visible inactivation, both, sphcn channels and HCN2 channels, display some kind of inactivation (7,22). Shin et al., (2004) suggested that HCN2 channels display a small prepulse-dependent inactivation which is camp dependent (22). However, this physiological prepulse-dependence of HCN2 is minor and cannot attribute to the very pronounced loss-of-function for hyperpolarizing voltage steps ranging up to -140 mv. From a mechanistic view, if the zipper mutants caused a full conversion of a `camp shift-type response, as in HCN channels (22), to a `camp inactivation-type response, as in sphcn channels (22), then one might indeed observe a strong reduction in current amplitudes. However, the `camp inactivation-type of sphcn channels is not apparent in recordings from intact Xenopus oocytes, due to the relatively high intracellular levels of camp (7). Thus, even a full conversion to a sphcn-like inactivation cannot explain the almost complete loss-of-functions which we have observed for some of the HCN leucine zipper mutants. In addition, camp dependent inactivation of sphcn channels occurs without a shift in the voltage dependence of activation (22), whereas the leucine zipper mutants have a strong shift in the voltage dependence of activation. We have studied several mutations at the lowermost zipper position of HCN2 (L343) and HCN1 (L290) (Fig. 6C). These data indicate that mutations at the lowermost zipper residues (L343 or L290) primarily cause a reduction of I max (Fig. 6C), similar as for the mutations introduced at the three other leucine zipper positions (Figs. 2, 3 and 4). However, we have observed a gain-offunction on current amplitude for arginine and glutamine mutations at position L343 which seem to be specific for HCN2. Most importantly, the reason for the difference on current magnitude for arginine and glutamine mutations at the lowermost S5 leucine zipper site of HCN1 and HCN2 might not be `found on our pore homology model. It is the leading hypothesis that camp-induced tetramerization of the C-terminal domains of HCN channels removes the tonic inhibition of the pore gate by the C-linker/CNBD. In this context it is noteworthy, that HCN1 channels have a different gating than HCN2 channels, as HCN1 channels open at less hyperpolarized potentials and exhibit only a minor response to camp. Recently, these differences in HCN gating were explained by the
9 effect that the C-linker/CNBD of HCN1 channels already form tetramers at basal camp concentrations (23-24); meaning that HCN1 channels are already disinhibited by the C-linker/CNBD. The difference between the interaction of the CNBD/C-linker with the late S5 and S6 of HCN2 and HCN1 might explain the apparent discrepancy of arginine und glutamine mutations at the lowest leucine zipper site. The glutamine and arginine mutations might exclusively lead to a gain-of-function in HCN2 by a disinhibition of the tonic block by the CNBD/C-linker, while HCN1 channels are already disinhibited. As for both HCN isoforms gain-of-functions or loss-of-functions were observed by glutamine and arginine mutations, the lowest zipper residue is a critical determinant for HCN1 and HCN2 channels to determine open-probability and the instantaneous current component. HCN channels expressed in heterologous systems give rise to two different currents, a slowly activating current I h and an instantaneous voltage independent current (25). The amplitude of both is correlated and both currents are modulated by changes in intracellular chloride and elevation of camp (25). Nevertheless, the two components of HCN channel currents were previously proposed to flow through two distinct channel populations (26). We found that mutations in the upper part of the leucine zipper drastically alter the voltage-dependent state of HCN channels and that mutations in the lower part of the leucine zipper modulate the instantaneous voltage independent open state. We therefore support the idea that HCN channels have two different open states and that both open states are critically dependent on the leucine zipper. Summarizing, we have observed that the leucine zipper plays a crucial role for HCN2 and HCN1 channel gating. The residues of the leucine zipper interact with the S6 segment and they are important molecular determinants for regulating the current amplitudes, voltage dependence and the closed state of hyperpolarization-activated cation channels. 9
10 REFERENCES 1. DiFrancesco, D. (1993) Pacemaker mechanisms in cardiac tissue. Annu Rev Physiol 55, Wahl-Schott, C., and Biel, M. (2009) HCN channels: structure, cellular regulation and physiological function. Cell Mol Life Sci 66, Santoro, B., Grant, S. G., Bartsch, D., and Kandel, E. R. (1997) Interactive cloning with the SH3 domain of N-src identifies a new brain specific ion channel protein, with homology to eag and cyclic nucleotide-gated channels. Proc Natl Acad Sci U S A 94, Ludwig, A., Zong, X., Jeglitsch, M., Hofmann, F., and Biel, M. (1998) A family of hyperpolarization-activated mammalian cation channels. Nature 393, Chen, J., Mitcheson, J. S., Lin, M., and Sanguinetti, M. C. (2000) Functional roles of charged residues in the putative voltage sensor of the HCN2 pacemaker channel. J Biol Chem 275, Chen, J., Mitcheson, J. S., Tristani-Firouzi, M., Lin, M., and Sanguinetti, M. C. (2001) The S4-S5 linker couples voltage sensing and activation of pacemaker channels. Proc Natl Acad Sci U S A 98, Männikkö, R., Elinder, F., and Larsson, H. P. (2002) Voltage-sensing mechanism is conserved among ion channels gated by opposite voltages. Nature 419, Tristani-Firouzi, M., Chen, J., and Sanguinetti, M. C. (2002) Interactions between S4-S5 linker and S6 transmembrane domain modulate gating of HERG K + channels. J Biol Chem 277, Decher, N., Chen, J., and Sanguinetti, M. C. (2004) Voltage-dependent gating of hyperpolarization-activated, cyclic nucleotide-gated pacemaker channels: molecular coupling between the S4-S5 and C-linkers. J Biol Chem 279, Landschulz, W. H., Johnson, P. F., and McKnight, S. L. (1988) The leucine zipper: a hypothetical structure common to a new class of DNA binding proteins. Science 240, McCormack, K., Tanouye, M. A., Iverson, L. E., Lin, J. W., Ramaswami, M., McCormack, T., Campanelli, J. T., Mathew, M. K., and Rudy, B. (1991) A role for hydrophobic residues in the voltage-dependent gating of Shaker K + channels. Proc Natl Acad Sci U S A 88, Syme, C. A., Hamilton, K. L., Jones, H. M., Gerlach, A. C., Giltinan, L., Papworth, G. D., Watkins, S. C., Bradbury, N. A., and Devor, D. C. (2003) Trafficking of the Ca 2+ -activated K + channel, hik1, is dependent upon a C-terminal leucine zipper. J Biol Chem 278, Hulme, J. T., Lin, T. W., Westenbroek, R. E., Scheuer, T., and Catterall, W. A. (2003) Betaadrenergic regulation requires direct anchoring of PKA to cardiac CaV1.2 channels via a leucine zipper interaction with A kinase-anchoring protein 15. Proc Natl Acad Sci U S A 100, Hulme, J. T., Ahn, M., Hauschka, S. D., Scheuer, T., and Catterall, W. A. (2002) A novel leucine zipper targets AKAP15 and cyclic AMP-dependent protein kinase to the C terminus of the skeletal muscle Ca 2+ channel and modulates its function. J Biol Chem 277, Zhong, H., Molday, L. L., Molday, R. S., and Yau, K. W. (2002) The heteromeric cyclic nucleotide-gated channel adopts a 3A:1B stoichiometry. Nature 420, Zerangue, N., Schwappach, B., Jan, Y. N., and Jan, L. Y. (1999) A new ER trafficking signal regulates the subunit stoichiometry of plasma membrane K ATP channels. Neuron 22, Sali, A., and Blundell, T. L. (1993) Comparative protein modelling by satisfaction of spatial restraints. J Mol Biol 234, Clayton, G. M., Altieri, S., Heginbotham, L., Unger, V. M., and Morais-Cabral, J. H. (2008) Structure of the transmembrane regions of a bacterial cyclic nucleotide-regulated channel. Proc Natl Acad Sci U S A 105, Long, S. B., Campbell, E. B., and Mackinnon, R. (2005) Crystal structure of a mammalian voltage-dependent Shaker family K + channel. Science 309,
11 20. Lu, Z., Klem, A. M., and Ramu, Y. (2002) Coupling between voltage sensors and activation gate in voltage-gated K + channels. J Gen Physiol 120, Sanguinetti, M. C., and Xu, Q. P. (1999) Mutations of the S4-S5 linker alter activation properties of HERG potassium channels expressed in Xenopus oocytes. J Physiol 514 ( Pt 3), Shin, K. S., Maertens, C., Proenza, C., Rothberg, B. S., and Yellen, G. (2004) Inactivation in HCN channels results from reclosure of the activation gate: desensitization to voltage. Neuron 41, Chow, S. S., Van Petegem, F., and Accili, E. A. (2012) Energetics of cyclic AMP binding to HCN channel C terminus reveal negative cooperativity. J Biol Chem 287, Lolicato, M., Nardini, M., Gazzarrini, S., Moller, S., Bertinetti, D., Herberg, F. W., Bolognesi, M., Martin, H., Fasolini, M., Bertrand, J. A., Arrigoni, C., Thiel, G., and Moroni, A. (2011) Tetramerization dynamics of C-terminal domain underlies isoform-specific camp gating in hyperpolarization-activated cyclic nucleotide-gated channels. J Biol Chem 286, Proenza, C., Angoli, D., Agranovich, E., Macri, V., and Accili, E. A. (2002) Pacemaker channels produce an instantaneous current. J Biol Chem 277, Proenza, C., and Yellen, G. (2006) Distinct populations of HCN pacemaker channels produce voltage-dependent and voltage-independent currents. J Gen Physiol 127, Acknowledgements- We thank Oxana Nowak and Maren Eisenbarth for a helping hand in molecular biology. We thank Michael Sanguinetti for HCN expression constructs. FOOTNOTES *This work was supported by grants DE1482-2/1 and DE1482-3/2 of the Deutsche Forschungsgemeinschaft (DFG) to N.D.. 1 To whom correspondence should be addressed: Niels Decher, PhD, Philipps University Marburg, Institute for Physiology and Pathophysiology, Vegetative Physiology, Deutschhausstrasse 1-2, D Marburg, Germany; Fax: (+49) ; decher@staff.uni-marburg.de 2 Structural Bioinformatics and Computational Biochemistry Unit, Department of Biochemistry, University of Oxford, South Parks Road, Oxford OX1 3QU, UK FIGURE LEGENDS FIGURE 1. Leucine zipper in HCN channels. A, The positions of the four leucine residues of the zipper motif in HCN channels and the closely related sphcn channel are highlighted in bold with boxes. The leucine zipper is not present in CNG and Kv channels. The numbers below the sequence provide the position of the leucine residues in mouse HCN2. B, Cartoon of a HCN2 channel indicating the position of the leucine zipper residues in the S5 segment. FIGURE 2. Electrophysiological properties of leucine zipper alanine mutants. A, Representative current traces for wild-type HCN2 and alanine mutants recorded during 2 s pulses from a holding potential of -30 mv to potentials ranging from -30 to -140 mv in 10 mv decrements. Tail currents were measured at -130 mv. The dotted line represents zero current. The arrow indicates the instantaneous current component. B, Relative currents measured at the end of a 2 s activating pulse to -130 mv, recorded two days after injection. Currents were normalized to wild-type HCN2 amplitude of every recording day. Wild-type HCN2 channel currents were 7.2 ± 0.4 µa (n = 46). The mean currents of L343A, L350, L357 and L364A were 2.5 ± 0.1 µa (n = 42), 1.3 ± 0.1 µa (n = 42), 0.8 ± 0.1 µa (n = 39) and 0.6 ± 0.1 µa (n = 36), respectively. C, Western blot of HCN2 channel proteins expressed in Xenopus oocytes after injection of the same amount of wild-type or mutant HCN2 crnas. HCN channels with an extracellular HA-epitope were detected by an HA-antibody and a peroxidase-linked secondary antibody. D, Relative surface expression determined by single cell luminometry. Surface expression was normalized to wild-type surface expression (0 ± 8). E, 11
12 Activation curves for wild-type HCN2 and alanine mutants of the leucine zipper. Relative currents were fitted to a Boltzmann equation. The V 1/2, slope factor and min-po values are given in Table 1. For L364A currents were normalized to wild-type HCN2 currents at -130 mv. FIGURE 3. Electrophysiological properties of double and triple alanine mutants in the leucine zipper of HCN2. A, Representative current traces for wild-type and double/triple alanine mutants recorded during 2 s pulses from a holding potential of -30 mv to potentials ranging from -30 to -140 mv in 10 mv decrements. Tail currents were measured at -130 mv. The dotted line represents zero current. B, Relative currents measured at the end of a 2 s activating pulse to -130 mv, recorded two days after injection. Currents were normalized to wild-type HCN2 amplitude of every recording day. Mean currents were: wild-type HCN2 7.2 ± 0.4 µa (n = 46), L343A/L350A 2.5 ± 0.3 µa (n = 14), L343A/L357A 1.2 ± 0.1 µa (n = 10), L350A/L357A 0.4 ± 0.1 µa (n = 20), L343A/L350A/L357A 0.4 ± 0.1 µa (n = 9). C, Relative surface expression determined by single cell luminometry after injection of equal amounts of crna. Surface expression was normalized to wild-type surface expression (0 ± 0.18). Inset displays the western blot to show total HCN2 expression for wild-type and double/triple alanine mutants in Xenopus oocytes, after injection of equal amounts of crna. The western blot was performed as described above. D, Activation curves for wild-type and mutant HCN2 channels. Relative currents were fitted to a Boltzmann equation. The V 1/2, slope factor and min-po values are given in Table 1. For L343A/L357A, L350A/L357A and L343A/L350A/L357A currents were normalized to wild-type amplitude at -130 mv. FIGURE 4. Electrophysiological properties of arginine and glutamine mutations in the leucine zipper of HCN2. A and F, Representative current traces for wild-type HCN2, glutamine (A) and arginine (F) mutants recorded during 2 s pulses from a holding potential of -30 mv to potentials ranging from -30 to -140 mv in 10 mv decrements. Tail currents were measured at -130 mv. The dotted line represents zero current. The arrow indicates the instantaneous current component. B and G, Relative currents of glutamine (B) and arginine (G) mutants measured at the end of a 2 s activating pulse to -130 mv, recorded two days after injection. Currents were normalized to wild-type HCN2 amplitude of every recording day. Current amplitudes were as follows: wild-type HCN2 7.2 ± 0.4 µa (n = 46), L343Q 43.5 ± 4.4 µa (n = 13), L350Q 0.2 ± 2 µa (n = 22), L357Q 0.3 ± 4 µa (n = 22) and L364Q 0.6 ± 0.1 µa (n = 24), L343R 14.8 ± 1.4 µa (n = 16), L350R 0.3 ± 0.1 µa (n = 24), L357R 0.5 ± 0.1 µa (n = 24) and 1.3 ± 0.2 µa (n = 23). C and H, Western blot of HCN2 channels expressed in Xenopus oocytes, after injection of equal amounts of crna encoding glutamine (C) and arginine (H) HCN2 mutants of the leucine zipper. Proteins with an introduced extracellular HA-tag were detected by an HA-antibody with a luminometric western blot. D and I, Relative surface expression of glutamine (D) and arginine (I) mutants determined by single cell luminometry. Surface expression was normalized to wild-type surface expression (0 ± 8). E and J, Activation curves for wild-type HCN2 glutamine (E) and arginine (J) mutants. Relative currents were fitted to a Boltzmann equation. The V 1/2, slope factor and min-po values are given in Table 1. For L357Q, L364Q, L357R and L364R currents were normalized to wild-type HCN2 currents at -130 mv. FIGURE 5. A closed state pore homology model predicts S6 segment residues to interact with the S5 segment leucine zipper. A, A closed state homology model of HCN2, based on the MlotiK1 crystal structure. Depicted in yellow are the residues of the leucine zipper and in magenta the interacting residues of the opposing S6 segment. B, Representative current traces of alanine mutations at S6 segment residues which were predicted to interact with the leucine zipper. The arrow indicates the instantaneous current component. C, Current amplitudes of alanine mutants in the leucine zipper of Figure 2 were compared to the opposing S6 segment residues, recorded two days after injection and analyzed at -130 mv. Currents were normalized to wild-type HCN2 amplitude of every recording day. Wild-type current amplitude was 8.0 ± 1.3 µa (n = 33) and mutant current amplitudes were: L416A 0.8 ± 0.1 µa (n = 39), V423A 2.2 ± 0.3 µa (n = 36), M430A 3.3 ± 0.4 µa (n = 36) and L438A 3.4 ± 12
13 0.3 µa (n = 31). The reduction in current amplitudes of the leucine zipper mutants correlates with the reduction in current amplitudes of the corresponding S6 segment mutants. FIGURE 6. Electrophysiological properties of HCN1 leucine zipper mutants. A, Representative current traces for HCN1 leucine zipper alanine mutants recorded during 2 s pulses from a holding potential of -30 mv to potentials ranging from -30 to -140 mv in 10 mv decrements. Tail currents were measured at -130 mv. The dotted line represents zero current. The arrow indicates the instantaneous current component. B, Currents measured at the end of a 2 s activating pulse to -130 mv, recorded two days after injection. Currents were normalized to wild-type HCN1 of every recording day. The mean HCN1 amplitude was 13.3 ± 1.6 µa (n = 39). The mean current amplitudes of the mutants L290Q, L297Q, L304Q and L311Q were, 0.5 ± 0.1 µa (n = 24), 0.4 ± 0.1 µa (n = 24), 1.9 ± 0.3 µa (n = 15) and ± 0.1 µa (n = 15), respectively. The V 1/2, slope factor and min-po values are provided in Table 2. C, Relative current amplitudes of mutations at position L343 of HCN2 and at position L290 of HCN1. Currents were measured at the end of a 2 s activating pulse to -130 mv, recorded two days after injection. Currents were normalized to the respective wild-type amplitudes of every recording day. FIGURE 7. Mutations at the lowermost leucine zipper residue and the interacting S6 residue lead to instantaneous currents in HCN1 and HCN2. A, Representative HCN1 currents of L290 mutations with an increased instantaneous current, see grey arrows at grey traces at -110 mv, to see the fraction of instantaneous current. The right panel illustrates the relative instantaneous current at -110 mv. B, Representative HCN2 currents of L343 and L438 mutations, displaying an increased relative instantaneous current, see arrows at grey traces at -110 mv, to see the fraction of instantaneous current. Right panel illustrates the relative instantaneous current at -110 mv. FIGURE 8. Fully-activated current-voltage relationships reveal no changes in ion selectivity for instantaneously open HCN2 and HCN1 mutants. A, Fully-activated current-voltage relationships for wild-type HCN2 and the S6 mutant L438R. For the representative current traces only the voltage steps around the reversal potential (-0 to -60 mv) are illustrated. The inset illustrates the reversal potentials for wild-type HCN2 and L438A. B, Fully-activated current-voltage relationships for wildtype HCN1 and the S5 leucine zipper mutant L290R. For the representative current traces only the voltage steps around the reversal potential (0 to -60 mv) are illustrated. The inset illustrates the reversal potentials for wild-type HCN1 and L290Q. Grey arrows in (A) and (B) indicate the fraction of instantaneous current. FIGURE 9. The leucine zipper stabilizes a salt-bridge of the S4-S5 linker and the C-linker, which is only present in the closed state of the channel. A, Closed state pore homology model of HCN2 based on the MlotiK1 crystal structure, with leucine zipper residues (yellow) and corresponding S6 segment sites (magenta). B, Depicted is an open state homology model of HCN2 based on the rkv1.2 crystal structure. The same color code as in (A) was used. R339 is illustrated in brown and D443 in pink. C, A single subunit of the closed state homology model of HCN2 and D, open state homology model, using the same color code as in (A and B). 13
14 TABLES Table 1: Properties of HCN2 leucine zipper mutations min-p o V 1/2 in mv k n HCN2 wild-type 53 ± ± ± L343A ± ± ± L350A 82 ± 05 * ± ± L357A 99 ± 06 ns ± ± L364A 59 ± 01 ns < -140 n.a 3 L343A/L350A ± ± 0.7 ns 6.8 ± L343A/L357A 43 ± 10 ns < -140 n.a 12 L350A/L357A 20 ± 02 < -140 n.a 8 L343A/L350A/L357A 29 ± 01 < -140 n.a 6 L343R ± ± ± L357R 33 ± 01 ** < -140 n.a 4 L364R 71 ± 01 ns < -140 n.a 6 L343Q 88 ± 06 ns ± 0.3 ns 6.2 ± L357Q 18 ± 01 < -140 n.a 4 L364Q 30 ± 01 ** < -140 n.a 9 min-p o = minimum open probability defined as the minimum value of relative tail current, V 1/2 = voltage required for half-maximal activation, k = slope of current voltage relationship, n.a. = not applicable. 14
15 Table 2: Properties of HCN1 leucine zipper mutations min-p o V 1/2 in mv k n HCN1 wild-type ± ± ± L290Q ± ± 2.3 ns 8.8 ± L304Q 94 ± 06 < -140 mv n.a 8 L311Q 67 ± 04 < -140 mv n.a 5 L290R ± 04 < -140 mv n.a 6 min-p o = minimum open probability defined as the minimum value of relative tail current, V 1/2 = voltage required for half-maximal activation, k = slope of current voltage relationship, n.a = not applicable. 15

16 Figure 1 A LxxYxxxLxxYxxxLxxYxxxLxxYxxx B mhcn1 ASAVVRIFNLIGMMLLLCHWDGCLQFLVPLLQDFPPDCWVSLNE 324 mhcn2 ASAVMRICNLISMMLLLCHWDGCL Q FLVPML Q DFPRNCWVSING 377 hhcn2 ASAVMRICNLISMMLLLCHWDGCLQFLVPMLQDFPRNCWVSING 404 mhcn3 ASAVVRIFNLIGMMLLLCHWDGCLQFLVPMLQDFPPDCWVSINH 287 mhcn4 ASAVVRIVNLIGMMLLLCHWDGCLQFLVPMLQDFPDDCWVSINN 455 sphcn ANAVIRICNLVCMMLLIGHWNGCLQYLVPMLQEYPDQSWVAING 405 mcnga1 YPNIFRISNLVMYIVIIIHWNACVYYSISKAIGFGNDTWVYPDV 329 mcnga2 YPNIFRISNLVLYILVIIHWNACIYYAISKSIGFGVDTWVYPNI 314 mkv1.5 LGLLIFFLFIGVILFSSAVYFAEADNQGSHFSSIPDAFWWAVVT 466 mkv4.3 LGFLLFSLTMAIIIFATVMFYAEKGSSASKFTSIPASFWYTIVT
17 Figure 2 A HCN2 B 0 L343A L357A C 2 μa 500 ms L350A L364A E Relative curren nt L343A L350A L357A L364A D Relative ligh ht units 1.2 n.s. n.s. n.s. n.s L343A L350A L357A L364A Normaliz zed current Voltage(mV) L343A L350A L357A L364A -20
18 Figure 3 A 1 μa HCN2 500 ms L343A/L350A L343A/L357A L350A/L357A B nt Relative curre L343A/L350A/L357A L343A/L350A L3 L343A/L357A L350A/L357A L343A/L350A/L357A C D 100 kd its Relative light uni 1.2 n.s. n.s L343A/L350A L3 L343A/L357A L350A/L357A L343A/L350A/L357A Normalized current L343A/L350A L343A/L357A L350A/L357A L343A/L350A/L357A Voltage (mv)
19 Figure 4 A HCN2 B C E 2 μa 500 ms L343Q L350Q L357Q L364Q Relative current Q 0Q 7Q 4Q L343Q L350Q L357Q L364Q D Relative light units n.s. 1.2 n.s. n.s. * L343Q3Q L350Q0Q L357Q7Q L364Q4Q Normalized current Voltage(mV) -40 L343Q L357Q L364Q -20 F 2 μa HCN2 500 ms L343R L350R L357R L364R G t Relative current L L L343R L350R L357R L364R L L H I Relative light units L n.s. n.s. n.s. n.s. L L343R L350R L357R L364R L L J Normalized curren nt Voltage(mV) L343R L357R L364R


20 Figure 5 A B L416A V423A 2 μa 500 ms M430A L438A L364 L416 L357 V423 L350 M430 L343 L438 L364 L416 V423 L357 S6 L350 M430 S5 L343 L438 S4-S5 C L364A L357A L350A L343A ** ** S5 0.5 S6 ** * ** 0.5 Relative current L416A V423A M430A L438A
21 Figure 6 1 μa A HCN1 500 ms L290Q L297Q L304Q L311Q B urrent Relative c 0.5 HCN1 L290Q L297Q L304Q L311Q C urrent Relative c control HCN2 L343A L343D L343E L343R L343Q L290R L290Q HCN1
22 Figure 7 A 1 μa HCN1 500 ms L290Q I Inst B µa 2µA ms 500 ms HCN2 L343A L343R I Inst L290R 0.2 L438A 0.2 HCN1 L290Q L290R L438R L343A L343R L438A L438R
23 Figure 8 A HCN2 2 μa A HCN2 L438R B HCN1 2 μa A 2 μa 500ms HCN1 L290R Voltage (mv) Voltage (mv) 500ms 500ms HCN2 L438R 1 μa 500ms Relative current Reversal potential HCN2 L438R HCN1 L290R Relative current Reversal potential HCN1 L290R












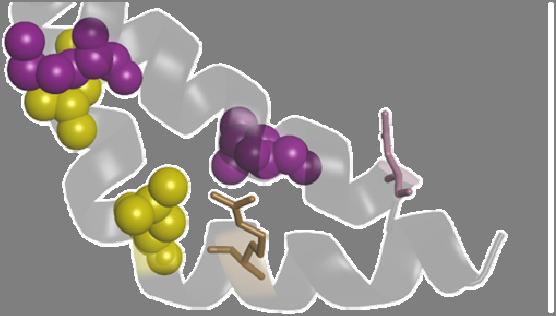


24 Figure 9 closed A D C closed open L364 L357 L350 M430 V423 L416 L438 R339 S5 S6 S4-S5 L364 L357 B L343 L416 V423 L364 L357 L416 V423 D443 open L364 L357 M430 V423 L416 R339 L350 M430 L343 L438 R339 D443 L350 M430 L343 L438 L364 L357 V423 L416 R339 D443 L343 M430 R339 D443 L438 L350 L343 L438 D443 L350
Transfer of ion binding site from ether-à-go-go to Shaker: Mg 2+ binds to resting state to modulate channel opening
 A r t i c l e Transfer of ion binding site from ether-à-go-go to Shaker: Mg 2+ binds to resting state to modulate channel opening Meng-chin A. Lin, 1 Jeff Abramson, 1 and Diane M. Papazian 1,2,3 1 Department
A r t i c l e Transfer of ion binding site from ether-à-go-go to Shaker: Mg 2+ binds to resting state to modulate channel opening Meng-chin A. Lin, 1 Jeff Abramson, 1 and Diane M. Papazian 1,2,3 1 Department
Module Membrane Biogenesis and Transport Lecture 15 Ion Channels Dale Sanders
 Module 0220502 Membrane Biogenesis and Transport Lecture 15 Ion Channels Dale Sanders 9 March 2009 Aims: By the end of the lecture you should understand The principles behind the patch clamp technique;
Module 0220502 Membrane Biogenesis and Transport Lecture 15 Ion Channels Dale Sanders 9 March 2009 Aims: By the end of the lecture you should understand The principles behind the patch clamp technique;
Ion Channel Structure and Function (part 1)
") Ion Channel Structure and Function (part 1) The most important properties of an ion channel Intrinsic properties of the channel (Selectivity and Mode of Gating) + Location Physiological Function Types
Ion Channel Structure and Function (part 1) The most important properties of an ion channel Intrinsic properties of the channel (Selectivity and Mode of Gating) + Location Physiological Function Types
Potassium channel gating and structure!
 Reading: Potassium channel gating and structure Hille (3rd ed.) chapts 10, 13, 17 Doyle et al. The Structure of the Potassium Channel: Molecular Basis of K1 Conduction and Selectivity. Science 280:70-77
Reading: Potassium channel gating and structure Hille (3rd ed.) chapts 10, 13, 17 Doyle et al. The Structure of the Potassium Channel: Molecular Basis of K1 Conduction and Selectivity. Science 280:70-77
Perspective Coupled Movements in Voltage-gated Ion Channels
 Perspective Coupled Movements in Voltage-gated Ion Channels Richard Horn Department of Physiology, Jefferson Medical College, Philadelphia, PA 19107 Ion channels, like most other proteins, are designed
Perspective Coupled Movements in Voltage-gated Ion Channels Richard Horn Department of Physiology, Jefferson Medical College, Philadelphia, PA 19107 Ion channels, like most other proteins, are designed
How is the heart rate regulated in the sinoatrial node? Another piece to the puzzle
 Published Online: 16 August, 2010 Supp Info: http://doi.org/10.1085/jgp.201010506 Downloaded from jgp.rupress.org on July 1, 2018 C o m m e n t a r y How is the heart rate regulated in the sinoatrial node?
Published Online: 16 August, 2010 Supp Info: http://doi.org/10.1085/jgp.201010506 Downloaded from jgp.rupress.org on July 1, 2018 C o m m e n t a r y How is the heart rate regulated in the sinoatrial node?
Transport of glucose across epithelial cells: a. Gluc/Na cotransport; b. Gluc transporter Alberts
 Figure 7 a. Secondary transporters make up the largest subfamily of transport proteins. TAGI 2000. Nature 408, 796 1. Na+- or H+-coupled cotransporters - Secondary active transport 2/7-02 Energy released
Figure 7 a. Secondary transporters make up the largest subfamily of transport proteins. TAGI 2000. Nature 408, 796 1. Na+- or H+-coupled cotransporters - Secondary active transport 2/7-02 Energy released
Homology models of the tetramerization domain of six eukaryotic voltage-gated potassium channels Kv1.1-Kv1.6
 Homology models of the tetramerization domain of six eukaryotic voltage-gated potassium channels Kv1.1-Kv1.6 Hsuan-Liang Liu* and Chin-Wen Chen Department of Chemical Engineering and Graduate Institute
Homology models of the tetramerization domain of six eukaryotic voltage-gated potassium channels Kv1.1-Kv1.6 Hsuan-Liang Liu* and Chin-Wen Chen Department of Chemical Engineering and Graduate Institute
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11054 Supplementary Fig. 1 Sequence alignment of Na v Rh with NaChBac, Na v Ab, and eukaryotic Na v and Ca v homologs. Secondary structural elements of Na v Rh are indicated above the
doi:10.1038/nature11054 Supplementary Fig. 1 Sequence alignment of Na v Rh with NaChBac, Na v Ab, and eukaryotic Na v and Ca v homologs. Secondary structural elements of Na v Rh are indicated above the
Supplementary Figure 1 Structure of the Orai channel. (a) The hexameric Drosophila Orai channel structure derived from crystallography 1 comprises
 The hexameric Drosophila Orai channel structure derived from crystallography 1 comprises") Supplementary Figure 1 Structure of the Orai channel. (a) The hexameric Drosophila Orai channel structure derived from crystallography 1 comprises six Orai subunits, each with identical amino acid sequences
Supplementary Figure 1 Structure of the Orai channel. (a) The hexameric Drosophila Orai channel structure derived from crystallography 1 comprises six Orai subunits, each with identical amino acid sequences
S3-S4 Linker Length Modulates the Relaxed State of a Voltage-Gated Potassium Channel
 2312 Biophysical Journal Volume 105 November 2013 2312 2322 S3-S4 Linker Length Modulates the Relaxed State of a Voltage-Gated Potassium Channel Michael F. Priest, Jérôme J. Lacroix, Carlos A. Villalba-Galea,
2312 Biophysical Journal Volume 105 November 2013 2312 2322 S3-S4 Linker Length Modulates the Relaxed State of a Voltage-Gated Potassium Channel Michael F. Priest, Jérôme J. Lacroix, Carlos A. Villalba-Galea,
7.06 Cell Biology EXAM #3 April 21, 2005
 7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
7.06 Cell Biology EXAM #3 April 21, 2005 This is an open book exam, and you are allowed access to books, a calculator, and notes but not computers or any other types of electronic devices. Please write
Binding Site in Eag Voltage Sensor Accommodates a Variety of Ions and is Accessible in Closed Channel
 3110 Biophysical Journal Volume 87 November 2004 3110 3121 Binding Site in Eag Voltage Sensor Accommodates a Variety of Ions and is Accessible in Closed Channel William R. Silverman, John P. A. Bannister,
3110 Biophysical Journal Volume 87 November 2004 3110 3121 Binding Site in Eag Voltage Sensor Accommodates a Variety of Ions and is Accessible in Closed Channel William R. Silverman, John P. A. Bannister,
Neuroscience 201A Exam Key, October 7, 2014
 Neuroscience 201A Exam Key, October 7, 2014 Question #1 7.5 pts Consider a spherical neuron with a diameter of 20 µm and a resting potential of -70 mv. If the net negativity on the inside of the cell (all
Neuroscience 201A Exam Key, October 7, 2014 Question #1 7.5 pts Consider a spherical neuron with a diameter of 20 µm and a resting potential of -70 mv. If the net negativity on the inside of the cell (all
Sriharsha Vemana, Shilpi Pandey, and H. Peter Larsson. Neurological Sciences Institute. Oregon Health & Science University. 505 NW 185 th Avenue
 Articles in PresS. Am J Physiol Cell Physiol (June 25, 2008). doi:10.1152/ajpcell.00154.2008 1 Intracellular Mg 2+ is a voltage dependent pore blocker of HCN channels Sriharsha Vemana, Shilpi Pandey, and
Articles in PresS. Am J Physiol Cell Physiol (June 25, 2008). doi:10.1152/ajpcell.00154.2008 1 Intracellular Mg 2+ is a voltage dependent pore blocker of HCN channels Sriharsha Vemana, Shilpi Pandey, and
Membrane Protein Channels
 Membrane Protein Channels Potassium ions queuing up in the potassium channel Pumps: 1000 s -1 Channels: 1000000 s -1 Pumps & Channels The lipid bilayer of biological membranes is intrinsically impermeable
Membrane Protein Channels Potassium ions queuing up in the potassium channel Pumps: 1000 s -1 Channels: 1000000 s -1 Pumps & Channels The lipid bilayer of biological membranes is intrinsically impermeable
Cardiac cell-cell Communication Part 1 Alonso P. Moreno D.Sc. CVRTI, Cardiology
 Bioengineering 6003 Cellular Electrophysiology and Biophysics Cardiac cell-cell Communication Part 1 Alonso P. Moreno D.Sc. CVRTI, Cardiology moreno@cvrti.utah.edu November 2010 poster Physiological Relevance
Bioengineering 6003 Cellular Electrophysiology and Biophysics Cardiac cell-cell Communication Part 1 Alonso P. Moreno D.Sc. CVRTI, Cardiology moreno@cvrti.utah.edu November 2010 poster Physiological Relevance
Supporting Material for. Microscopic origin of gating current fluctuations in a potassium channel voltage sensor
 Supporting Material for Microscopic origin of gating current fluctuations in a potassium channel voltage sensor J. Alfredo Freites, * Eric V. Schow, * Stephen H. White, and Douglas J. Tobias * * Department
Supporting Material for Microscopic origin of gating current fluctuations in a potassium channel voltage sensor J. Alfredo Freites, * Eric V. Schow, * Stephen H. White, and Douglas J. Tobias * * Department
LESSON 2.2 WORKBOOK How do our axons transmit electrical signals?
 LESSON 2.2 WORKBOOK How do our axons transmit electrical signals? This lesson introduces you to the action potential, which is the process by which axons signal electrically. In this lesson you will learn
LESSON 2.2 WORKBOOK How do our axons transmit electrical signals? This lesson introduces you to the action potential, which is the process by which axons signal electrically. In this lesson you will learn
The Potassium Ion Channel: Rahmat Muhammad
 The Potassium Ion Channel: 1952-1998 1998 Rahmat Muhammad Ions: Cell volume regulation Electrical impulse formation (e.g. sodium, potassium) Lipid membrane: the dielectric barrier Pro: compartmentalization
The Potassium Ion Channel: 1952-1998 1998 Rahmat Muhammad Ions: Cell volume regulation Electrical impulse formation (e.g. sodium, potassium) Lipid membrane: the dielectric barrier Pro: compartmentalization
Supplementary figure 1 Application of tmfret in LeuT. (a) To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a
 To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a") Supplementary figure 1 Application of tmfret in LeuT. (a) To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a series of tmfret-pairs comprised of single cysteine mutants
Supplementary figure 1 Application of tmfret in LeuT. (a) To assess the feasibility of using tmfret for distance-dependent measurements in LeuT, a series of tmfret-pairs comprised of single cysteine mutants
SUPPLEMENTARY INFORMATION. doi: /nature07461
 Figure S1 Electrophysiology. a ph-activation of. Two-electrode voltage clamp recordings of Xenopus oocytes expressing in comparison to waterinjected oocytes. Currents were recorded at 40 mv. The ph of
Figure S1 Electrophysiology. a ph-activation of. Two-electrode voltage clamp recordings of Xenopus oocytes expressing in comparison to waterinjected oocytes. Currents were recorded at 40 mv. The ph of
V m = the Value of the Na Battery Plus the Voltage Drop Across g Na. I Na is Isolated By Blocking I K. and g K
 VoltageGated Ion Channels and the Action Potential VoltageGated Ion Channels and the Action Potential jdk3 Principles of Neural Science, chaps 8&9 The Action Potential Generation Conduction VoltageGated
VoltageGated Ion Channels and the Action Potential VoltageGated Ion Channels and the Action Potential jdk3 Principles of Neural Science, chaps 8&9 The Action Potential Generation Conduction VoltageGated
Mutations in the S4 domain of a pacemaker channel alter its voltage dependence
 FEBS 23956 FEBS Letters 479 (2000) 35^40 Mutations in the S4 domain of a pacemaker channel alter its voltage dependence L. Vaca a;b; *, J. Stieber b, X. Zong b, A. Ludwig b, F. Hofmann b, M. Biel b;c a
FEBS 23956 FEBS Letters 479 (2000) 35^40 Mutations in the S4 domain of a pacemaker channel alter its voltage dependence L. Vaca a;b; *, J. Stieber b, X. Zong b, A. Ludwig b, F. Hofmann b, M. Biel b;c a
Building a Homology Model of the Transmembrane Domain of the Human Glycine α-1 Receptor
 Building a Homology Model of the Transmembrane Domain of the Human Glycine α-1 Receptor Presented by Stephanie Lee Research Mentor: Dr. Rob Coalson Glycine Alpha 1 Receptor (GlyRa1) Member of the superfamily
Building a Homology Model of the Transmembrane Domain of the Human Glycine α-1 Receptor Presented by Stephanie Lee Research Mentor: Dr. Rob Coalson Glycine Alpha 1 Receptor (GlyRa1) Member of the superfamily
The principle of gating charge movement in a voltage-dependent K 1 channel
 The principle of gating charge movement in a voltage-dependent K 1 channel Youxing Jiang*, Vanessa Ruta, Jiayun Chen, Alice Lee & Roderick MacKinnon Howard Hughes Medical Institute, Laboratory of Molecular
The principle of gating charge movement in a voltage-dependent K 1 channel Youxing Jiang*, Vanessa Ruta, Jiayun Chen, Alice Lee & Roderick MacKinnon Howard Hughes Medical Institute, Laboratory of Molecular
Equivalent Circuit of the Membrane Connected to the Voltage Clamp. I mon. For Large Depolarizations, Both I Na and I K Are Activated
 VoltageGated Ion Channels and the Action Potential jdk3 Principles of Neural Science, chaps 8&9 VoltageGated Ion Channels and the Action Potential The Action Potential Generation Conduction VoltageGated
VoltageGated Ion Channels and the Action Potential jdk3 Principles of Neural Science, chaps 8&9 VoltageGated Ion Channels and the Action Potential The Action Potential Generation Conduction VoltageGated
Inhibition of S532C by MTSET at intracellular ph 6.8 indicates accessibility in the closed
 Supplementary Text Inhibition of S532C by MTSET at intracellular ph 6.8 indicates accessibility in the closed state It is difficult to examine accessibility of cysteine-substituted mutants in the fully
Supplementary Text Inhibition of S532C by MTSET at intracellular ph 6.8 indicates accessibility in the closed state It is difficult to examine accessibility of cysteine-substituted mutants in the fully
Channels can be activated by ligand-binding (chemical), voltage change, or mechanical changes such as stretch.
, voltage change, or mechanical changes such as stretch.") 1. Describe the basic structure of an ion channel. Name 3 ways a channel can be "activated," and describe what occurs upon activation. What are some ways a channel can decide what is allowed to pass through?
1. Describe the basic structure of an ion channel. Name 3 ways a channel can be "activated," and describe what occurs upon activation. What are some ways a channel can decide what is allowed to pass through?
Name: TF: Section Time: LS1a ICE 5. Practice ICE Version B
 Name: TF: Section Time: LS1a ICE 5 Practice ICE Version B 1. (8 points) In addition to ion channels, certain small molecules can modulate membrane potential. a. (4 points) DNP ( 2,4-dinitrophenol ), as
Name: TF: Section Time: LS1a ICE 5 Practice ICE Version B 1. (8 points) In addition to ion channels, certain small molecules can modulate membrane potential. a. (4 points) DNP ( 2,4-dinitrophenol ), as
RNA Polymerase I Contains a TFIIF-Related DNA-Binding Subcomplex
 Molecular Cell, Volume 39 Supplemental Information RNA Polymerase I Contains a TFIIFRelated DNABinding Subcomplex Sebastian R. Geiger, Kristina Lorenzen, Amelie Schreieck, Patrizia Hanecker, Dirk Kostrewa,
Molecular Cell, Volume 39 Supplemental Information RNA Polymerase I Contains a TFIIFRelated DNABinding Subcomplex Sebastian R. Geiger, Kristina Lorenzen, Amelie Schreieck, Patrizia Hanecker, Dirk Kostrewa,
Interpreting and evaluating biological NMR in the literature. Worksheet 1
 Interpreting and evaluating biological NMR in the literature Worksheet 1 1D NMR spectra Application of RF pulses of specified lengths and frequencies can make certain nuclei detectable We can selectively
Interpreting and evaluating biological NMR in the literature Worksheet 1 1D NMR spectra Application of RF pulses of specified lengths and frequencies can make certain nuclei detectable We can selectively
Introduction to Comparative Protein Modeling. Chapter 4 Part I
 Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Structure and Function of Voltage-Gated Sodium Channels at Atomic Resolution
 Structure and Function of Voltage-Gated Sodium Channels at Atomic Resolution Royal Society of Chemistry Cambridge UK, March 2013 William A. Catterall, Department of Pharmacology, University of Washington
Structure and Function of Voltage-Gated Sodium Channels at Atomic Resolution Royal Society of Chemistry Cambridge UK, March 2013 William A. Catterall, Department of Pharmacology, University of Washington
SUPPLEMENTARY INFORMATION
 Supplementary Table 1: Amplitudes of three current levels. Level 0 (pa) Level 1 (pa) Level 2 (pa) TrkA- TrkH WT 200 K 0.01 ± 0.01 9.5 ± 0.01 18.7 ± 0.03 200 Na * 0.001 ± 0.01 3.9 ± 0.01 12.5 ± 0.03 200
Supplementary Table 1: Amplitudes of three current levels. Level 0 (pa) Level 1 (pa) Level 2 (pa) TrkA- TrkH WT 200 K 0.01 ± 0.01 9.5 ± 0.01 18.7 ± 0.03 200 Na * 0.001 ± 0.01 3.9 ± 0.01 12.5 ± 0.03 200
External Barium Affects the Gating of KCNQ1 Potassium Channels and Produces a Pore Block via Two Discrete Sites
 External Barium Affects the Gating of KCNQ1 Potassium Channels and Produces a Pore Block via Two Discrete Sites Gilad Gibor, Daniel Yakubovich, Asher Peretz, and Bernard Attali Department of Physiology
External Barium Affects the Gating of KCNQ1 Potassium Channels and Produces a Pore Block via Two Discrete Sites Gilad Gibor, Daniel Yakubovich, Asher Peretz, and Bernard Attali Department of Physiology
Overview of ion channel proteins. What do ion channels do? Three important points:
 Overview of ion channel proteins Protein Structure Membrane proteins & channels Specific channels Several hundred distinct types Organization Evolution We need to consider 1. Structure 2. Functions 3.
Overview of ion channel proteins Protein Structure Membrane proteins & channels Specific channels Several hundred distinct types Organization Evolution We need to consider 1. Structure 2. Functions 3.
Influence of permeating ions on potassium channel block by external tetraethylammonium
 4414 Journal of Physiology (1995), 486.2, pp.267-272 267 Influence of permeating ions on potassium channel block by external tetraethylammonium Stephen R. Ikeda* and Stephen J. Korn t *Department of Pharmacology
4414 Journal of Physiology (1995), 486.2, pp.267-272 267 Influence of permeating ions on potassium channel block by external tetraethylammonium Stephen R. Ikeda* and Stephen J. Korn t *Department of Pharmacology
cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody
 Based on Monoclonal Anti-cGMP Antibody") (FOR RESEARCH USE ONLY. DO NOT USE IT IN CLINICAL DIAGNOSIS!) cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody Catalog No: E-EL-DS02 96T This manual must be read attentively and
(FOR RESEARCH USE ONLY. DO NOT USE IT IN CLINICAL DIAGNOSIS!) cgmp ELISA Kit (Direct Competitive) Based on Monoclonal Anti-cGMP Antibody Catalog No: E-EL-DS02 96T This manual must be read attentively and
Signal Transduction Phosphorylation Protein kinases. Misfolding diseases. Protein Engineering Lysozyme variants
 Signal Transduction Phosphorylation Protein kinases Misfolding diseases Protein Engineering Lysozyme variants Cells and Signals Regulation The cell must be able to respond to stimuli Cellular activities
Signal Transduction Phosphorylation Protein kinases Misfolding diseases Protein Engineering Lysozyme variants Cells and Signals Regulation The cell must be able to respond to stimuli Cellular activities
Physiology Unit 2. MEMBRANE POTENTIALS and SYNAPSES
 Physiology Unit 2 MEMBRANE POTENTIALS and SYNAPSES In Physiology Today Ohm s Law I = V/R Ohm s law: the current through a conductor between two points is directly proportional to the voltage across the
Physiology Unit 2 MEMBRANE POTENTIALS and SYNAPSES In Physiology Today Ohm s Law I = V/R Ohm s law: the current through a conductor between two points is directly proportional to the voltage across the
Introduction to electrophysiology. Dr. Tóth András
 Introduction to electrophysiology Dr. Tóth András Topics Transmembran transport Donnan equilibrium Resting potential Ion channels Local and action potentials Intra- and extracellular propagation of the
Introduction to electrophysiology Dr. Tóth András Topics Transmembran transport Donnan equilibrium Resting potential Ion channels Local and action potentials Intra- and extracellular propagation of the
Neurobiology Biomed 509 Ion Channels
 eurobiology Biomed 509 Ion hannels References: Luo 2.4, 2.4 2., (3.20 3.2), M 2.8 I. ores vs. carriers In order for hydrophilic ions to transit the hydrophobic membrane, they need to have a specific pathway
eurobiology Biomed 509 Ion hannels References: Luo 2.4, 2.4 2., (3.20 3.2), M 2.8 I. ores vs. carriers In order for hydrophilic ions to transit the hydrophobic membrane, they need to have a specific pathway
Participation of the S4 voltage sensor in the Mg 2 -dependent activation of large conductance (BK) K channels
 K channels") Participation of the S4 voltage sensor in the Mg 2 -dependent activation of large conductance (BK) K channels Lei Hu*, Jingyi Shi*, Zhongming Ma, Gayathri Krishnamoorthy*, Fred Sieling*, Guangping Zhang,
Participation of the S4 voltage sensor in the Mg 2 -dependent activation of large conductance (BK) K channels Lei Hu*, Jingyi Shi*, Zhongming Ma, Gayathri Krishnamoorthy*, Fred Sieling*, Guangping Zhang,
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11085 Supplementary Tables: Supplementary Table 1. Summary of crystallographic and structure refinement data Structure BRIL-NOP receptor Data collection Number of crystals 23 Space group
doi:10.1038/nature11085 Supplementary Tables: Supplementary Table 1. Summary of crystallographic and structure refinement data Structure BRIL-NOP receptor Data collection Number of crystals 23 Space group
Differential polyamine sensitivity in inwardly rectifying Kir2 potassium channels
 J Physiol 571.2 (2006) pp 287 302 287 Differential polyamine sensitivity in inwardly rectifying Kir2 potassium channels Brian K. Panama and Anatoli N. Lopatin University of Michigan, Department of Molecular
J Physiol 571.2 (2006) pp 287 302 287 Differential polyamine sensitivity in inwardly rectifying Kir2 potassium channels Brian K. Panama and Anatoli N. Lopatin University of Michigan, Department of Molecular
The Lipid Protein Interface of a Shaker K Channel
 The Lipid Protein Interface of a Shaker K Channel Kwang Hee Hong and Christopher Miller From the Department of Biochemistry, Howard Hughes Medical Institute, Brandeis University, Waltham, Massachusetts
The Lipid Protein Interface of a Shaker K Channel Kwang Hee Hong and Christopher Miller From the Department of Biochemistry, Howard Hughes Medical Institute, Brandeis University, Waltham, Massachusetts
Secondary Structure. Bioch/BIMS 503 Lecture 2. Structure and Function of Proteins. Further Reading. Φ, Ψ angles alone determine protein structure
 Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
S3-S4 Linker Modulation of Voltage Sensor Relaxation in Voltage Gated Potassium Channels
 Virginia Commonwealth University VCU Scholars Compass Theses and Dissertations Graduate School 2013 S3-S4 Linker Modulation of Voltage Sensor Relaxation in Voltage Gated Potassium Channels W. Everett Fox
Virginia Commonwealth University VCU Scholars Compass Theses and Dissertations Graduate School 2013 S3-S4 Linker Modulation of Voltage Sensor Relaxation in Voltage Gated Potassium Channels W. Everett Fox
Conversation between Voltage Sensors and Gates of Ion Channels
 Copyright 2000 by the American Chemical Society Volume 39, Number 51 December 26, 2000 Current Topics Conversation between Voltage Sensors and Gates of Ion Channels Richard Horn* Department of Physiology,
Copyright 2000 by the American Chemical Society Volume 39, Number 51 December 26, 2000 Current Topics Conversation between Voltage Sensors and Gates of Ion Channels Richard Horn* Department of Physiology,
Direct detection of antibodies in blood plasma using bioluminescent
 Supplementary information Direct detection of antibodies in blood plasma using bioluminescent sensor proteins and a smartphone. Remco Arts, Ilona den Hartog, Stefan Zijlema, Vito Thijssen, Stan van der
Supplementary information Direct detection of antibodies in blood plasma using bioluminescent sensor proteins and a smartphone. Remco Arts, Ilona den Hartog, Stefan Zijlema, Vito Thijssen, Stan van der
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/3/4/e1600663/dc1 Supplementary Materials for A dynamic hydrophobic core orchestrates allostery in protein kinases Jonggul Kim, Lalima G. Ahuja, Fa-An Chao, Youlin
advances.sciencemag.org/cgi/content/full/3/4/e1600663/dc1 Supplementary Materials for A dynamic hydrophobic core orchestrates allostery in protein kinases Jonggul Kim, Lalima G. Ahuja, Fa-An Chao, Youlin
Voltage-dependent gating of ion channels Prof. Keith S. Elmslie
 Voltage-Dependent Gating of Ion Channels Department of Pharmacology A.T. Still University Kirksville College of Osteopathic Medicine 1 The action potential (AP) E Na Threshold The N eur onal AP Sensory
Voltage-Dependent Gating of Ion Channels Department of Pharmacology A.T. Still University Kirksville College of Osteopathic Medicine 1 The action potential (AP) E Na Threshold The N eur onal AP Sensory
Nature Structural and Molecular Biology: doi: /nsmb.2783
 Supplementary Figure 1: Crystallized chimera construct (mhv1cc). (a) Sequence alignment between mhv1cc and other VSDs. These sequences (mhv1cc, Kv1.2 Kv2.1; shaker family voltage gated potassium channel
Supplementary Figure 1: Crystallized chimera construct (mhv1cc). (a) Sequence alignment between mhv1cc and other VSDs. These sequences (mhv1cc, Kv1.2 Kv2.1; shaker family voltage gated potassium channel
Introduction to electrophysiology 1. Dr. Tóth András
 Introduction to electrophysiology 1. Dr. Tóth András Topics Transmembran transport Donnan equilibrium Resting potential Ion channels Local and action potentials Intra- and extracellular propagation of
Introduction to electrophysiology 1. Dr. Tóth András Topics Transmembran transport Donnan equilibrium Resting potential Ion channels Local and action potentials Intra- and extracellular propagation of
Potential, Structure, and Excitability of Giant Axon Membrane
 Potential, Structure, and Excitability of Giant Axon Membrane T. NARAHASHI From the Laboratory of Applied Entomology, Faculty of Agriculture, University of Tokyo, Tokyo, Japan Intracellular perfusion of
Potential, Structure, and Excitability of Giant Axon Membrane T. NARAHASHI From the Laboratory of Applied Entomology, Faculty of Agriculture, University of Tokyo, Tokyo, Japan Intracellular perfusion of
ClC-1 inhibition by low ph and ATP INHIBITION OF SKELETAL MUSCLE ClC-1 CHLORIDE CHANNELS BY LOW INTRACELLULAR ph AND ATP
 JBC Papers in Press. Published on August 10, 2007 as Manuscript M703259200 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m703259200 INHIBITION OF SKELETAL MUSCLE ClC-1 CHLORIDE CHANNELS
JBC Papers in Press. Published on August 10, 2007 as Manuscript M703259200 The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.m703259200 INHIBITION OF SKELETAL MUSCLE ClC-1 CHLORIDE CHANNELS
Supplemental Information. The Mitochondrial Fission Receptor MiD51. Requires ADP as a Cofactor
 Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Structure, Volume 22 Supplemental Information The Mitochondrial Fission Receptor MiD51 Requires ADP as a Cofactor Oliver C. Losón, Raymond Liu, Michael E. Rome, Shuxia Meng, Jens T. Kaiser, Shu-ou Shan,
Supplementary Figure S1. MscS orientation in spheroplasts and liposomes (a) Current-voltage relationship for wild-type MscS expressed in E.
 Current-voltage relationship for wild-type MscS expressed in E.") a b c Supplementary Figure S1. MscS orientation in spheroplasts and liposomes (a) Current-voltage relationship for wild-type MscS expressed in E. coli giant spheroplasts (MJF465) and reconstituted into
a b c Supplementary Figure S1. MscS orientation in spheroplasts and liposomes (a) Current-voltage relationship for wild-type MscS expressed in E. coli giant spheroplasts (MJF465) and reconstituted into
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature17991 Supplementary Discussion Structural comparison with E. coli EmrE The DMT superfamily includes a wide variety of transporters with 4-10 TM segments 1. Since the subfamilies of the
doi:10.1038/nature17991 Supplementary Discussion Structural comparison with E. coli EmrE The DMT superfamily includes a wide variety of transporters with 4-10 TM segments 1. Since the subfamilies of the
Patrick: An Introduction to Medicinal Chemistry 5e Chapter 04
 01) Which of the following statements is not true about receptors? a. Most receptors are proteins situated inside the cell. b. Receptors contain a hollow or cleft on their surface which is known as a binding
01) Which of the following statements is not true about receptors? a. Most receptors are proteins situated inside the cell. b. Receptors contain a hollow or cleft on their surface which is known as a binding
Closing In on the Resting State of the Shaker K + Channel
 Article Closing In on the Resting State of the Shaker K + Channel Medha M. Pathak, 1,8,9 Vladimir Yarov-Yarovoy, 4,8 Gautam Agarwal, 2 Benoît Roux, 6 Patrick Barth, 5 Susy Kohout, 3 Francesco Tombola,
Article Closing In on the Resting State of the Shaker K + Channel Medha M. Pathak, 1,8,9 Vladimir Yarov-Yarovoy, 4,8 Gautam Agarwal, 2 Benoît Roux, 6 Patrick Barth, 5 Susy Kohout, 3 Francesco Tombola,
A conserved arginine near the filter of Kir1.1 controls Rb/K selectivity
 Research paper Channels 4:3, 1-12; May/June 2010; 2010 Landes Bioscience Research Paper A conserved arginine near the filter of Kir1.1 controls Rb/K selectivity This manuscript has been published online,
Research paper Channels 4:3, 1-12; May/June 2010; 2010 Landes Bioscience Research Paper A conserved arginine near the filter of Kir1.1 controls Rb/K selectivity This manuscript has been published online,
SECOND PUBLIC EXAMINATION. Honour School of Physics Part C: 4 Year Course. Honour School of Physics and Philosophy Part C C7: BIOLOGICAL PHYSICS
 2757 SECOND PUBLIC EXAMINATION Honour School of Physics Part C: 4 Year Course Honour School of Physics and Philosophy Part C C7: BIOLOGICAL PHYSICS TRINITY TERM 2013 Monday, 17 June, 2.30 pm 5.45 pm 15
2757 SECOND PUBLIC EXAMINATION Honour School of Physics Part C: 4 Year Course Honour School of Physics and Philosophy Part C C7: BIOLOGICAL PHYSICS TRINITY TERM 2013 Monday, 17 June, 2.30 pm 5.45 pm 15
Membrane Potentials, Action Potentials, and Synaptic Transmission. Membrane Potential
 Cl Cl - - + K + K+ K + K Cl - 2/2/15 Membrane Potentials, Action Potentials, and Synaptic Transmission Core Curriculum II Spring 2015 Membrane Potential Example 1: K +, Cl - equally permeant no charge
Cl Cl - - + K + K+ K + K Cl - 2/2/15 Membrane Potentials, Action Potentials, and Synaptic Transmission Core Curriculum II Spring 2015 Membrane Potential Example 1: K +, Cl - equally permeant no charge
I. MEMBRANE POTENTIALS
 I. MEMBRANE POTENTIALS Background to Nerve Impulses We have all heard that nerve impulses are electrical impulses. Stimuli at one end of a nerve cell are communicated to the far end of the nerve cell through
I. MEMBRANE POTENTIALS Background to Nerve Impulses We have all heard that nerve impulses are electrical impulses. Stimuli at one end of a nerve cell are communicated to the far end of the nerve cell through
Editorial. What is the true resting potential of small cells? Jean-Marc Dubois
 Gen. Physiol. Biophys. (2000), 19, 3 7 3 Editorial What is the true resting potential of small cells? Jean-Marc Dubois In order to understand almost anything, it is necessary to first obtain a measurement
Gen. Physiol. Biophys. (2000), 19, 3 7 3 Editorial What is the true resting potential of small cells? Jean-Marc Dubois In order to understand almost anything, it is necessary to first obtain a measurement
A) at equilibrium B) endergonic C) endothermic D) exergonic E) exothermic.
 at equilibrium B) endergonic C) endothermic D) exergonic E) exothermic.") CHEM 2770: Elements of Biochemistry Mid Term EXAMINATION VERSION A Date: October 29, 2014 Instructor: H. Perreault Location: 172 Schultz Time: 4 or 6 pm. Duration: 1 hour Instructions Please mark the Answer
CHEM 2770: Elements of Biochemistry Mid Term EXAMINATION VERSION A Date: October 29, 2014 Instructor: H. Perreault Location: 172 Schultz Time: 4 or 6 pm. Duration: 1 hour Instructions Please mark the Answer
A Single Histidine Residue Determines the ph Sensitivity of the. Pacemaker Channel HCN2
 JBC Papers in Press. Published on November 28, 2000 as Manuscript M010326200 A Single Histidine Residue Determines the ph Sensitivity of the Pacemaker Channel HCN2 Xiangang Zong*, Juliane Stieber*, Andreas
JBC Papers in Press. Published on November 28, 2000 as Manuscript M010326200 A Single Histidine Residue Determines the ph Sensitivity of the Pacemaker Channel HCN2 Xiangang Zong*, Juliane Stieber*, Andreas
Visual pigments. Neuroscience, Biochemistry Dr. Mamoun Ahram Third year, 2019
 Visual pigments Neuroscience, Biochemistry Dr. Mamoun Ahram Third year, 2019 References Webvision: The Organization of the Retina and Visual System (http://www.ncbi.nlm.nih.gov/books/nbk11522/#a 127) The
Visual pigments Neuroscience, Biochemistry Dr. Mamoun Ahram Third year, 2019 References Webvision: The Organization of the Retina and Visual System (http://www.ncbi.nlm.nih.gov/books/nbk11522/#a 127) The
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles
 Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Illegitimate translation causes unexpected gene expression from on-target out-of-frame alleles created by CRISPR-Cas9 Shigeru Makino, Ryutaro Fukumura, Yoichi Gondo* Mutagenesis and Genomics Team, RIKEN
Extracellular Sodium Interacts with the HERG Channel at an Outer Pore Site
 Extracellular Sodium Interacts with the HERG Channel at an Outer Pore Site Franklin M. Mullins, 1 Svetlana Z. Stepanovic, 2 Reshma R. Desai, 1 Alfred L. George, Jr., 1,3 and Jeffrey R. Balser 1,2 1 Department
Extracellular Sodium Interacts with the HERG Channel at an Outer Pore Site Franklin M. Mullins, 1 Svetlana Z. Stepanovic, 2 Reshma R. Desai, 1 Alfred L. George, Jr., 1,3 and Jeffrey R. Balser 1,2 1 Department
SUPPLEMENTARY INFORMATION
 Figure S1. Secondary structure of CAP (in the camp 2 -bound state) 10. α-helices are shown as cylinders and β- strands as arrows. Labeling of secondary structure is indicated. CDB, DBD and the hinge are
Figure S1. Secondary structure of CAP (in the camp 2 -bound state) 10. α-helices are shown as cylinders and β- strands as arrows. Labeling of secondary structure is indicated. CDB, DBD and the hinge are
Ion Channels. 6 th December, Touqeer Ahmed PhD Atta-ur-Rahman School of Applied Biosciences National University of Sciences and Technology
 Ion Channels 6 th December, 2016 Touqeer Ahmed PhD Atta-ur-Rahman School of Applied Biosciences National University of Sciences and Technology Introduction The ion channels of nerve cells are optimally
Ion Channels 6 th December, 2016 Touqeer Ahmed PhD Atta-ur-Rahman School of Applied Biosciences National University of Sciences and Technology Introduction The ion channels of nerve cells are optimally
SUPPLEMENTARY INFORMATION
 www.nature.com/nature 1 Figure S1 Sequence alignment. a Structure based alignment of the plgic of E. chrysanthemi (ELIC), the acetylcholine binding protein from the snail Lymnea stagnalis (AchBP, PDB code
www.nature.com/nature 1 Figure S1 Sequence alignment. a Structure based alignment of the plgic of E. chrysanthemi (ELIC), the acetylcholine binding protein from the snail Lymnea stagnalis (AchBP, PDB code
a Brownian model of the voltage sensor
 Simulation of gating currents of the Shaker K channel using a Brownian model of the voltage sensor Luigi Catacuzzeno * and Fabio Franciolini Department of Chemistry, Biology and Biotechnology University
Simulation of gating currents of the Shaker K channel using a Brownian model of the voltage sensor Luigi Catacuzzeno * and Fabio Franciolini Department of Chemistry, Biology and Biotechnology University
Introduction Principles of Signaling and Organization p. 3 Signaling in Simple Neuronal Circuits p. 4 Organization of the Retina p.
 Introduction Principles of Signaling and Organization p. 3 Signaling in Simple Neuronal Circuits p. 4 Organization of the Retina p. 5 Signaling in Nerve Cells p. 9 Cellular and Molecular Biology of Neurons
Introduction Principles of Signaling and Organization p. 3 Signaling in Simple Neuronal Circuits p. 4 Organization of the Retina p. 5 Signaling in Nerve Cells p. 9 Cellular and Molecular Biology of Neurons
Functional Crosstalk Between α6β4 Nicotinic. Acetylcholine Receptors and P2X Receptors
 96 Chapter 4 Functional Crosstalk Between α6β4 Nicotinic Acetylcholine Receptors and P2X Receptors 4.1 Introduction Nicotinic acetylcholine receptors (nachrs) and P2X receptors are ligandgated cation channels
96 Chapter 4 Functional Crosstalk Between α6β4 Nicotinic Acetylcholine Receptors and P2X Receptors 4.1 Introduction Nicotinic acetylcholine receptors (nachrs) and P2X receptors are ligandgated cation channels
Structural basis for gating charge movement in the voltage sensor of a sodium channel. 1 V.Y.-Y. and P.G.D. contributed equally to this work.
 Structural basis for gating charge movement in the voltage sensor of a sodium channel Vladimir Yarov-Yarovoy a,1,2, Paul G. DeCaen a,1,3, Ruth E. Westenbroek a, Chien-Yuan Pan a,4, Todd Scheuer a, David
Structural basis for gating charge movement in the voltage sensor of a sodium channel Vladimir Yarov-Yarovoy a,1,2, Paul G. DeCaen a,1,3, Ruth E. Westenbroek a, Chien-Yuan Pan a,4, Todd Scheuer a, David
Voltage-dependent gating characteristics of the K channel KAT1 depend on the N and C termini
 Proc. Natl. Acad. Sci. USA Vol. 94, pp. 3448 3453, April 1997 Plant Biology Voltage-dependent gating characteristics of the K channel KAT1 depend on the N and C termini IRENE MARTEN AND TOSHINORI HOSHI*
Proc. Natl. Acad. Sci. USA Vol. 94, pp. 3448 3453, April 1997 Plant Biology Voltage-dependent gating characteristics of the K channel KAT1 depend on the N and C termini IRENE MARTEN AND TOSHINORI HOSHI*
Characterization of cardiac I Ks channel gating using voltage clamp fluorometry. Jeremiah D. Osteen
 Characterization of cardiac I Ks channel gating using voltage clamp fluorometry Jeremiah D. Osteen Submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the Graduate
Characterization of cardiac I Ks channel gating using voltage clamp fluorometry Jeremiah D. Osteen Submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the Graduate
The Nervous System and the Sodium-Potassium Pump
 The Nervous System and the Sodium-Potassium Pump 1. Define the following terms: Ion: A Student Activity on Membrane Potentials Cation: Anion: Concentration gradient: Simple diffusion: Sodium-Potassium
The Nervous System and the Sodium-Potassium Pump 1. Define the following terms: Ion: A Student Activity on Membrane Potentials Cation: Anion: Concentration gradient: Simple diffusion: Sodium-Potassium
9.01 Introduction to Neuroscience Fall 2007
 MIT OpenCourseWare http://ocw.mit.edu 9.01 Introduction to Neuroscience Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms. 9.01 Recitation (R02)
MIT OpenCourseWare http://ocw.mit.edu 9.01 Introduction to Neuroscience Fall 2007 For information about citing these materials or our Terms of Use, visit: http://ocw.mit.edu/terms. 9.01 Recitation (R02)
Supporting Information
 Supporting Information SUPPLEMENTARY MATERIAL SI Methods Multiple Sequence Alignment of Voltage-Gated Sodium and Potassium Channels. Sequence alignment between VSDs of the Na V and K V channels is well
Supporting Information SUPPLEMENTARY MATERIAL SI Methods Multiple Sequence Alignment of Voltage-Gated Sodium and Potassium Channels. Sequence alignment between VSDs of the Na V and K V channels is well
Department of Physiology and Nora Eccles Harrison Cardiovascular Research and Training Institute, University of Utah, Salt Lake City, UT 84112, USA
 J Physiol 563.2 (2005) pp 359 368 359 Mutation of colocalized residues of the pore helix and transmembrane segments S5 and S6 disrupt deactivation and modify inactivation of KCNQ1 K + channels Guiscard
J Physiol 563.2 (2005) pp 359 368 359 Mutation of colocalized residues of the pore helix and transmembrane segments S5 and S6 disrupt deactivation and modify inactivation of KCNQ1 K + channels Guiscard
Supplementary Figure 1
 Supplementary Figure 1 Activation of P2X2 receptor channels in symmetric Na + solutions only modestly alters the intracellular ion concentration. a,b) ATP (30 µm) activated P2X2 receptor channel currents
Supplementary Figure 1 Activation of P2X2 receptor channels in symmetric Na + solutions only modestly alters the intracellular ion concentration. a,b) ATP (30 µm) activated P2X2 receptor channel currents
Phys498BIO; Prof. Paul Selvin Hw #9 Assigned Wed. 4/18/12: Due 4/25/08
 1. Ionic Movements Across a Permeable Membrane: The Nernst Potential. In class we showed that if a non-permeable membrane separates a solution with high [KCl] from a solution with low [KCl], the net charge
1. Ionic Movements Across a Permeable Membrane: The Nernst Potential. In class we showed that if a non-permeable membrane separates a solution with high [KCl] from a solution with low [KCl], the net charge
Muscle regulation and Actin Topics: Tropomyosin and Troponin, Actin Assembly, Actin-dependent Movement
 1 Muscle regulation and Actin Topics: Tropomyosin and Troponin, Actin Assembly, Actin-dependent Movement In the last lecture, we saw that a repeating alternation between chemical (ATP hydrolysis) and vectorial
1 Muscle regulation and Actin Topics: Tropomyosin and Troponin, Actin Assembly, Actin-dependent Movement In the last lecture, we saw that a repeating alternation between chemical (ATP hydrolysis) and vectorial
BSc and MSc Degree Examinations
 Examination Candidate Number: Desk Number: BSc and MSc Degree Examinations 2018-9 Department : BIOLOGY Title of Exam: Molecular Biology and Biochemistry Part I Time Allowed: 1 hour and 30 minutes Marking
Examination Candidate Number: Desk Number: BSc and MSc Degree Examinations 2018-9 Department : BIOLOGY Title of Exam: Molecular Biology and Biochemistry Part I Time Allowed: 1 hour and 30 minutes Marking
Lecture goals: Learning Objectives
 Title: Membrane Potential in Excitable Cells 1 Subtitle: Voltage-Gated Ion Channels and the basis of the Action Potential Diomedes E. Logothetis, Ph.D. Lecture goals: This first of two lectures will use
Title: Membrane Potential in Excitable Cells 1 Subtitle: Voltage-Gated Ion Channels and the basis of the Action Potential Diomedes E. Logothetis, Ph.D. Lecture goals: This first of two lectures will use
Neurophysiology. Danil Hammoudi.MD
 Neurophysiology Danil Hammoudi.MD ACTION POTENTIAL An action potential is a wave of electrical discharge that travels along the membrane of a cell. Action potentials are an essential feature of animal
Neurophysiology Danil Hammoudi.MD ACTION POTENTIAL An action potential is a wave of electrical discharge that travels along the membrane of a cell. Action potentials are an essential feature of animal
Modulation of Kir1.1 Inactivation by Extracellular Ca and Mg
 Biophysical Journal Volume 100 March 2011 1207 1215 1207 Modulation of Kir1.1 Inactivation by Extracellular Ca and Mg Henry Sackin, * Mikheil Nanazashvili, Hui Li, Lawrence G. Palmer, and Lei Yang Department
Biophysical Journal Volume 100 March 2011 1207 1215 1207 Modulation of Kir1.1 Inactivation by Extracellular Ca and Mg Henry Sackin, * Mikheil Nanazashvili, Hui Li, Lawrence G. Palmer, and Lei Yang Department
Chapter 1 subtitles Ion gradients
 CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 1 subtitles Ion gradients Introduction In this first chapter, I'll explain the basic knowledge required to understand the electrical signals generated
CELLULAR NEUROPHYSIOLOGY CONSTANCE HAMMOND Chapter 1 subtitles Ion gradients Introduction In this first chapter, I'll explain the basic knowledge required to understand the electrical signals generated
Supporting Information
 Supporting Information Mullins et al. 10.1073/pnas.0906781106 SI Text Detection of Calcium Binding by 45 Ca 2 Overlay. The 45 CaCl 2 (1 mci, 37 MBq) was obtained from NEN. The general method of 45 Ca 2
Supporting Information Mullins et al. 10.1073/pnas.0906781106 SI Text Detection of Calcium Binding by 45 Ca 2 Overlay. The 45 CaCl 2 (1 mci, 37 MBq) was obtained from NEN. The general method of 45 Ca 2
The NH 2 Terminus of RCK1 Domain Regulates Ca 2 -dependent BK Ca Channel Gating
 ARTICLE The NH 2 Terminus of RCK1 Domain Regulates Ca 2 -dependent BK Ca Channel Gating Gayathri Krishnamoorthy, 1 Jingyi Shi, 2,3 David Sept, 2,4 and Jianmin Cui 2,3 1 Department of Biomedical Engineering,
ARTICLE The NH 2 Terminus of RCK1 Domain Regulates Ca 2 -dependent BK Ca Channel Gating Gayathri Krishnamoorthy, 1 Jingyi Shi, 2,3 David Sept, 2,4 and Jianmin Cui 2,3 1 Department of Biomedical Engineering,
Coupling of Voltage Sensing to Channel Opening Reflects Intrasubunit Interactions in Kv Channels
 Coupling of Voltage Sensing to Channel Opening Reflects Intrasubunit Interactions in Kv Channels Alain J. Labro, Adam L. Raes, and Dirk J. Snyders Laboratory for Molecular Biophysics, Physiology, and Pharmacology,
Coupling of Voltage Sensing to Channel Opening Reflects Intrasubunit Interactions in Kv Channels Alain J. Labro, Adam L. Raes, and Dirk J. Snyders Laboratory for Molecular Biophysics, Physiology, and Pharmacology,
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Transmembrane Domains (TMDs) of ABC transporters
 of ABC transporters") Transmembrane Domains (TMDs) of ABC transporters Most ABC transporters contain heterodimeric TMDs (e.g. HisMQ, MalFG) TMDs show only limited sequence homology (high diversity) High degree of conservation
Transmembrane Domains (TMDs) of ABC transporters Most ABC transporters contain heterodimeric TMDs (e.g. HisMQ, MalFG) TMDs show only limited sequence homology (high diversity) High degree of conservation
An NH2-Terminal Multi-Basic RKR Motif Is Required for the ATP-Dependent Regulation of hik1
 Channels ISSN: 1933-6950 (Print) 1933-6969 (Online) Journal homepage: http://www.tandfonline.com/loi/kchl20 An NH2-Terminal Multi-Basic RKR Motif Is Required for the ATP-Dependent Regulation of hik1 Heather
Channels ISSN: 1933-6950 (Print) 1933-6969 (Online) Journal homepage: http://www.tandfonline.com/loi/kchl20 An NH2-Terminal Multi-Basic RKR Motif Is Required for the ATP-Dependent Regulation of hik1 Heather
Structure of the SPRY domain of human DDX1 helicase, a putative interaction platform within a DEAD-box protein
 Supporting information Volume 71 (2015) Supporting information for article: Structure of the SPRY domain of human DDX1 helicase, a putative interaction platform within a DEAD-box protein Julian Kellner
Supporting information Volume 71 (2015) Supporting information for article: Structure of the SPRY domain of human DDX1 helicase, a putative interaction platform within a DEAD-box protein Julian Kellner