BAM receptors regulate stem cell specification and organ development. Department of Molecular, Cellular and Developmental Biology, University of
|
|
|
- Barrie McCarthy
- 5 years ago
- Views:
Transcription
1 Genetics: Published Articles Ahead of Print, published on September 9, 2008 as /genetics BAM receptors regulate stem cell specification and organ development through complex interactions with CLAVATA signaling Brody J. DeYoung 1 and Steven E. Clark 2 Department of Molecular, Cellular and Developmental Biology, University of Michigan, Ann Arbor, MI Current address: Department of Biology, Indiana University, Bloomington, IN , bdeyoung@indiana.edu 2 Correspoinding author: Department of Molecular, Cellular and Developmental Biology, University of Michigan, 830 North University, Ann Arbor, MI clarks@umich.edu; phone ; fax
2 Running Head: Analysis of CLV-related receptors Key Words/Phrases: meristem, stem cell, organogenesis, differentiation, signal transduction Corresponding author: Steven E. Clark, Department of Molecular, Cellular and Developmental Biology, University of Michigan, 830 North University, Ann Arbor, MI , USA. phone ; fax
3 ABSTRACT The CLAVATA1 (CLV1) receptor kinase regulates stem cell specification at shoot and flower meristems of Arabidopsis. Most clv1 alleles are dominantnegative, and clv1 null alleles are weak in phenotype, suggesting additional receptors functioning in parallel. We have identified two such parallel receptors, BAM1 and BAM2. We show that the weak nature of the phenotype of clv1 null alleles is dependent on BAM activity, with bam clv mutants exhibiting severe defects in stem cell specification. Furthermore, BAM activity in the meristem depends on CLV2, which is required in part for CLV1 function. In addition, clv1 mutants enhance many of the Bam - organ phenotypes, indicating that, contrary to current understanding, CLV1 function is not specific to the meristem. CLV3 encodes a small, secreted peptide that acts as the ligand for CLV1. Mutations in clv3 lead to increased stem cell accumulation. Surprisingly, bam1 and bam2 mutants suppress the phenotype of clv3 mutants. We speculate that in addition to redundant function in the meristem center, BAM1 and BAM2 act to sequester CLV3-like ligands in the meristem flanks. 3
4 INTRODUCTION The vast majority of tissues and organs of the adult plant are formed postembryonically. To achieve continuous organ formation plants rely on the activity of stem cells located at meristems (BYRNE et al. 2003; CARLES and FLETCHER 2003; GROSS-HARDT and LAUX 2003). The shoot apical meristem, which produces the aerial portion of the plant, is composed of a central pool of stem cells surrounded by descendant cells that are directed towards differentiation. Division of stem cells results in daughter cells that maintain the central stem cell population as well as daughter cells that are localized more peripherally, where they perceive positional cues that drive their differentiation. Stem cell specification and organogenesis at the Arabidopsis shoot apical meristem is regulated by a receptor-kinase signaling system that includes the CLAVATA1 (CLV1), CLV2 and CLV3 gene products. CLV1 encodes a receptorkinase protein with 21 leucine-rich repeats (LRRs) in its predicted extracellular domain, a single pass transmembrane domain, and a cytoplasmic serine/threonine kinase domain (CLARK et al. 1997). CLV2 encodes a protein structurally similar to CLV1, however CLV2 has a very small predicted cytoplasmic domain with no known signaling motifs (JEONG et al. 1999). CLV1 accumulation is at least in part dependent on the presence of CLV2 (JEONG et al. 1999). Furthermore, CLV1 and CLV2 are hypothesized to form a heterodimer based on migration in both gel chromatography and non-reducing protein gel blots (TROTOCHAUD et al. 1999). 4
5 Several lines of evidence suggest that CLV3 is the ligand for CLV1. CLV3 is both secreted and appears to be subjected to proteolytic maturation, releasing a conserved polypetide, termed the CLE domain (for CLV3/ESR-related), from the C-terminus that is capable of altering plant development when added exogenously (FIERS et al. 2005; ITO et al. 2006; KONDO et al. 2006; NI and CLARK 2006). Null mutations in CLV3 are largely epistatic to mutations in CLV1 and CLV2, while CLV3 over-expression phenotypes are dependent on CLV1, suggesting that CLV3 functions upstream of CLV1 (BRAND et al. 2000; CLARK et al. 1995). In addition, CLV3 is required for formation of a large molecular mass complex containing CLV1 (TROTOCHAUD et al. 1999). CLV1 acts to limit the diffusion of CLV3 protein suggesting that CLV1 binds to CLV3, sequestering it (LENHARD and LAUX 2003). Recent work has shown that the CLE peptide is capable of binding to the CLV1 extracellular domain (OGAWA et al. 2008). Taken together these data suggest a model in which CLV1 and CLV2 form a receptor complex that is capable of activating specific signal transduction pathways upon perception of the CLE peptide derived from CLV3. CLV1, CLV2, and CLV3 function in a common genetic pathway to regulate the expression domain of the stem cell-promoting transcription factor, WUSCHEL (WUS) (BRAND et al. 2000; MAYER et al. 1998; SCHOOF et al. 2000). Mutations in CLV1, CLV2, or CLV3 result in an increased WUS expression domain and a larger meristem with extra stem cells (BRAND et al. 2000; CLARK et al. 1993; CLARK et al. 1995; KAYES and CLARK 1998; SCHOOF et al. 2000), while mutations in WUS result in reduction or elimination of the meristem (LAUX et al. 1996). The 5
6 related phosphatases POLTERGEIST (POL) and PLL1 are signaling intermediates acting downstream of CLV1, CLV2 and CLV3. POL/PLL1 act to maintain WUS expression within the meristem, while CLV signaling represses POL/PLL1 activity (SONG and CLARK 2005; SONG et al. 2006; YU et al. 2003; YU et al. 2000). The remaining genetic and biochemical mechanisms that link CLV1 activation and WUS expression are largely unknown. Several lines of evidence suggest the presence of additional receptors that function in parallel with CLV1. Most of the clv1 mutant alleles identified in mutagenic screens are dominant-negative missense alleles that exhibit intermediate to strong phenotypes, while clv1 null alleles exhibit quite weak phenotypes by comparison (DIéVART et al. 2003). In addition, clv3 null alleles exhibit strong meristem phenotypes and are epistatic to both null and dominantnegative alleles of clv1, demonstrating that CLV3 retains some function in the absence of CLV1 (CLARK et al. 1995). Candidates for CLV1-redundant receptors can be found among the related BAM receptor-kinases (BAM1, BAM2, and BAM3) (DEYOUNG et al. 2006). CLV1 and the BAM genes form a monophyletic group of Arabidopsis genes encoding related receptor-kinases, although the duplication giving rise to the BAM and CLV1 clades predates the split between monocots and dicots (DEYOUNG et al. 2006). Despite the related sequences of the these receptors, several functional differences exist between CLV1 and BAM receptors (DEYOUNG et al. 2006). While CLV1 promotes stem cell differentiation, BAM receptors are required for stem cell maintenance. In bam1 bam2 double mutants, a reduction in meristem 6
7 size is observed, while in bam1 bam2 bam3 triple mutants a frequent incidence of premature shoot and flower meristem termination is observed. A second key difference lies in the specificity of the receptors. clv1 mutants only exhibit phenotypes at the shoot and flower meristem, while all three BAM receptors regulate numerous aspects of Arabidopsis development, from leaf vascular patterning, to the proper development of all floral organs, especially the anthers and ovules (CLARK et al. 1993; DEYOUNG et al. 2006; HORD et al. 2006). Despite these differences, CLV1 and BAM receptors appear to have retained significant similarities in protein function, because broadly expressed CLV1 can fully rescue bam mutants while meristem-expressed BAM receptors can partially rescue clv1 mutants (DEYOUNG et al. 2006). Thus, on a genetic level, the differences between CLV1 and BAM function appear to be largely driven by expression differences. Consistent with this, BAM1 and BAM2 are expressed throughout the plant and exhibit relatively high levels of expression on the meristem flanks, while CLV1 is expressed primarily in the meristem center (CLARK 1997; DEYOUNG et al. 2006). To understand the relationships between CLV1, CLV2 and BAM receptors, especially within the meristem where they play apparently complimentary roles, we sought to assess the genetic interactions of BAM1 and BAM2 with mutant alleles of CLV1, CLV2, and CLV3. We show that bam and clv mutants display complex gene- and allele-specific interactions. bam mutants suppress clv3 phenotypes, have no effect on clv2 phenotypes, and enhance clv1 null phenotypes. Our evidence suggests that BAM proteins function similarly to 7
8 CLV1 on a biochemical level and, in fact, function in parallel with CLV1 in the meristem center, but carry out separate roles elsewhere within the plant. We propose that one of these roles is to sequester CLE peptide ligands on the flanks of the meristem, creating a buffer around the meristem that prevents these ligands from upsetting the delicate balance necessary for stem cell maintenance. 8
9 MATERIALS AND METHODS Plant growth (YU et al. 2003), SEM analysis (DIéVART et al. 2003), and phloroglucinol staining (PRIGGE et al. 2005) were performed as previously described unless otherwise noted. Genetic Analysis: clv1-1 bam1-1, clv1-4 bam1-1, clv1-7 bam1-1, clv1-11 bam1-1, clv2-1 bam1-1, clv3-2 bam1-1, clv1-1 bam2-1, clv1-4 bam2-1, clv1-7 bam2-1, clv1-11 bam2-1, clv2-1 bam2-1, clv3-2 bam2-1, clv1-7 bam1-3, clv1-11 bam1-3, clv1-7 bam2-3, and clv1-11 bam2-3 double mutant combinations were generated by crossing plants homozygous for the bam1-1, bam2-1, bam1-3, or bam2-3 allele to plants homozygous for the respective clv1, clv2, or clv3 mutant allele. Individual F 2 progeny were screened on the basis of phenotype to identify individuals homozygous for the respective clv1, clv2, or clv3 mutant alleles and, in the cases of bam1-1 and bam1-3, to identify lines homozygous for the er-1 allele. No segregation distortion was observed that might suggest clv homozygous mutants were not segregating in a 3:1 ratio. These plants were subsequently screened using a PCR strategy to identify individuals homozygous for bam1-1, bam2-1, bam1-3, or bam2-3, respectively as previously described (DEYOUNG et al. 2006). In some cases, progeny from F 2 plants heterozygous for bam1-1, bam2-1, bam1-3, or bam2-3 were screened in the F 3 generation to identify lines homozygous for bam1-1, bam2-1, bam1-3, or bam2-3, respectively. 9
10 clv1-1 bam1-1 bam2-1, clv1-4 bam1-1 bam2-1, clv1-7 bam1-1 bam2-1, clv1-11 bam1-1 bam2-1, clv2-1 bam1-1 bzm2-1, clv3-2 bam1-1 bam2-1 triple mutant combinations were generated by crossing clv bam1 double mutant plants to the respective clv bam2 mutant plants. For example, clv1-1 bam1-1 bam2-1 plants were generated by crossing clv1-1 bam1-1 plants to clv1-1 bam2-1 and screening the F 2 progeny of this cross to identify plants homozygous for both bam1-1 and bam2-1. Because the bam1-1 bam2-1 allele combination causes infertility, the bam1 bam2 clv1-1 plants used for subsequent analysis were identified from segregating progeny of sibling plants heterozygous at one BAM locus and homozygous mutant at the other, as above. Floral Phenotype Measurement: The mean number of carpels per flower was determined by averaging the number of carpels per flower from the first 10 flowers of each plant. Image Analysis: Photographic mages were captured with a Nikon Coolpix 995 digital camera. Brightness and contrast of some photographic and SEM images were adjusted with Adobe PHOTOSHOP CS. Accession Numbers: Sequence data from this article can be found in the EMBL/GenBank data libraries under accession numbers Q9SYQ8 ( AT1G75820) for CLV1, AAF02655 (AT1G65380) for CLV2, AAD27620 (AT2G27250) for CLV3, NM_ (AT5G65700) for BAM1, and NM_ (AT3G49670) for 10
11 BAM2. All bam alleles used in this study have been provided to the Arabidopsis Biological Resource Center ( 11
12 RESULTS clv3 phenotypes are suppressed by bam1 bam2 mutations: Our previous genetic studies have suggested that BAM and CLV proteins have retained significant biochemical redundancy, although the developmental role of BAM1 and BAM2 is apparently antagonistic to that of CLV1 in the meristem (DEYOUNG et al. 2006). We therefore undertook experiments to characterize the role of BAM receptors in the absence of specific CLV pathway components in an attempt to clarify the role of the BAM receptors within meristem development. In particular, it is critical to investigate the relationship(s) between BAM and CLV3, because the expression of these genes in adjacent cell populations raises the possibility that CLV3 acts as a ligand for the BAM receptors on the flanks of the meristem. To this end, bam1-1 and bam2-1 single and double mutants were generated in the clv3-2 null mutant background. In order to quantify the relative effect of these mutations on the flower meristem, we took advantage of previous observations that floral organ number, especially carpel number, reflects the relative size of the flower meristem and is a sensitive method of detecting change in flower meristem size (CLARK et al. 1993; SONG and CLARK 2005). When carpel number per flower was assessed in bam1 clv3 and bam2 clv3 plants, we observed a modest, but statistically significant reduction when compared to clv3 single mutant plants (Table 1). bam1 mutations lead to a larger absolute reduction in carpel number in the clv3-2 mutant background than bam2. bam1 12
13 bam2 clv3 triple mutants exhibited more obvious suppression of the Clv phenotype, with approximately 4.3 carpels per flower compared to clv3-2 single mutants which have approximately 6.7 carpels per flower (Table 1). The suppression of clv3 by bam mutations is consistent with effects of multiple bam mutants to reduce meristem size (DEYOUNG et al. 2006). clv2 phenotypes are unaltered by bam1 bam2: CLV2 function is necessary for normal CLV1 function (JEONG et al. 1999). Because CLV2 expression can be detected broadly within the meristem, as well as outside of the meristem, and because clv2 mutants exhibit several phenotypes during organ development, we have hypothesized that CLV2 interacts with multiple RLKs in addition to CLV1 (JEONG et al. 1999; KAYES and CLARK 1998). Considering these observations, CLV2 could conceivably act as a partner for the BAM receptors. To test this idea, bam1-1 and bam2-1 single and double mutants were combined with the clv2-1 null mutation. While bam1 clv2 and bam2 clv2 exhibited a modest but statistically significant suppression in carpel number compared to clv2 single mutants, the mean carpel number of bam1 bam clv2 triple mutant flowers was not significantly different from that of clv2-1 alone (Table 1). Thus, clv2 is unaltered bam1 bam2 double mutants, in direct contrast to clv3. This raises the possibility that CLV2 may be necessary for both CLV1 and BAM function, which would imply that the Clv2 phenotype is the combined result of the loss of both the meristem-inhibiting CLV1 pathway and the meristempromoting BAM pathway. 13
14 Synergistic meristem interactions of bam1, bam2 and clv1 null alleles: To determine the net effect of removing both BAM and CLV1 receptors from the meristem, bam1-1 and bam2-1 mutations were combined with clv1-7 or clv1-11 mutations. clv1-11 is a null allele containing a T-DNA insertion in the coding sequence for the LRR domain, while clv1-7 contains a nonsense mutation that results in truncation of a large portion of the kinase domain and appears phenotypically similar to clv1 null alleles (DIéVART et al. 2003). Unexpectedly, bam1 and bam2 alleles significantly enhanced the flower meristem defects in both the clv1-11 and clv1-7 backgrounds (Table 2). To test if these results could be attributed to linked mutations in the bam1-1 or bam2-1 background, we repeated these experiments with the bam1-3 and bam2-3 alleles. Again we observed a significant enhancement of the meristem defect (Table 2). For all combinations, bam1 alleles resulted in a stronger enhancement than bam2 alleles. Interestingly, bam1 caused a stronger suppression of clv3 than bam2 (Table 1). To assess the effect of removing all three receptor kinases on meristem development, bam1-1 bam2-1 clv1-11 and bam1-1 bam2-1 clv1-7 triple mutant plants were generated. In both cases the bam1 bam2 clv1 triple mutants exhibited rather extreme phenotypes, with the mean number of carpels per flower greater than 8.6 for the clv1-11 combination (Table 2). To determine if these unexpected interactions were specific to the flower meristem, we examined shoot apical meristem size and organization in clv1, 14
15 bam1 clv1, bam2 clv1, and bam1 bam2 clv1 triple mutant plants. In each case, bam mutations enhanced the shoot meristem defects of clv1-7 and clv1-11, in accordance with the flower meristem enhancement. Namely, bam1 clv1 were more enhanced compared to clv1 than bam2 clv1, and the bam1 bam2 clv1 triple mutant exhibited extreme shoot meristem defects (Figure 1). Indeed, the bam1 bam2 clv1 triple mutants exhibited novel phenotypes that are discussed below. These data are not consistent with a model in which BAM genes simply function in a separate pathway antagonistic to the CLV pathway. The observation that BAM genes are required for the weak phenotype of clv1 null alleles suggests that that the BAM and CLV loci interact in a complex manner and that BAM and CLV1 genes may act in a functionally redundant manner in the center of the meristem. Divergent interactions of clv1 dominant-negative alleles: The apparent functional overlap of the BAM and CLV1 receptors evident from the bam clv1 combinations could occur either within the same cells, or within separate cells. Overlap occurring within separate cells would suggest that BAM acts in a complicated fashion on the meristem periphery to regulate meristem development. Overlap within the same cells would suggest that although BAM expression is low at the center of the meristem (DEYOUNG et al. 2006), functional BAM receptors are still present and acting redundantly with CLV1 in these cells. To distinguish between these possibilities, we combined bam1 and bam2 mutations with the clv1 dominant-negative alleles clv1-1 and clv1-4. These 15
16 alleles contain missense mutations in the kinase domain and LRR domain, respectively, and result in phenotypes significantly more severe than clv1 null alleles (CLARK et al. 1997; DIéVART et al. 2003). We have hypothesized that clv1 dominant-negative alleles block the action of receptors kinases with functional overlap. If the overlapping function of BAM receptors with CLV1 occurs in separate cells, we would expect similar interactions of bam mutants with clv1 null and dominant-negative alleles. If the functional overlap occurs within the same cells, clv1 dominant-negative alleles might partially block BAM1 and/or BAM2 function. When the bam clv1-1 double mutants were assayed, we observed that neither the bam1 clv1-1 nor the bam2 clv1-1 double mutants exhibited a stronger phenotype than clv1-1 alone (Table 2). This indicates that the genetic interactions of bam1 and bam2 with clv1-1 are different from those with the clv1 null alleles. When determining the genetic interactions with clv1-4, we observed a different result: both bam1 and bam2 were capable of enhancing the clv1-4 allele. Thus, the nature of the dominant-negative effect in the kinase domain missense allele (clv1-1) may be different from the LRR domain missense allele (clv1-4). The LRR domain dominant-negative alleles of clv1 are invariably more severe phenotypically than the kinase domain dominant-negative alleles (DIéVART et al. 2003), providing additional evidence of functional differences in dominant-negative action between LRR and kinase mutants. Neither clv1-1 nor clv1-4 exhibit the most severe clv mutant phenotypes, and CLV3 still functions in these mutant backgrounds (CLARK et al. 1995). 16
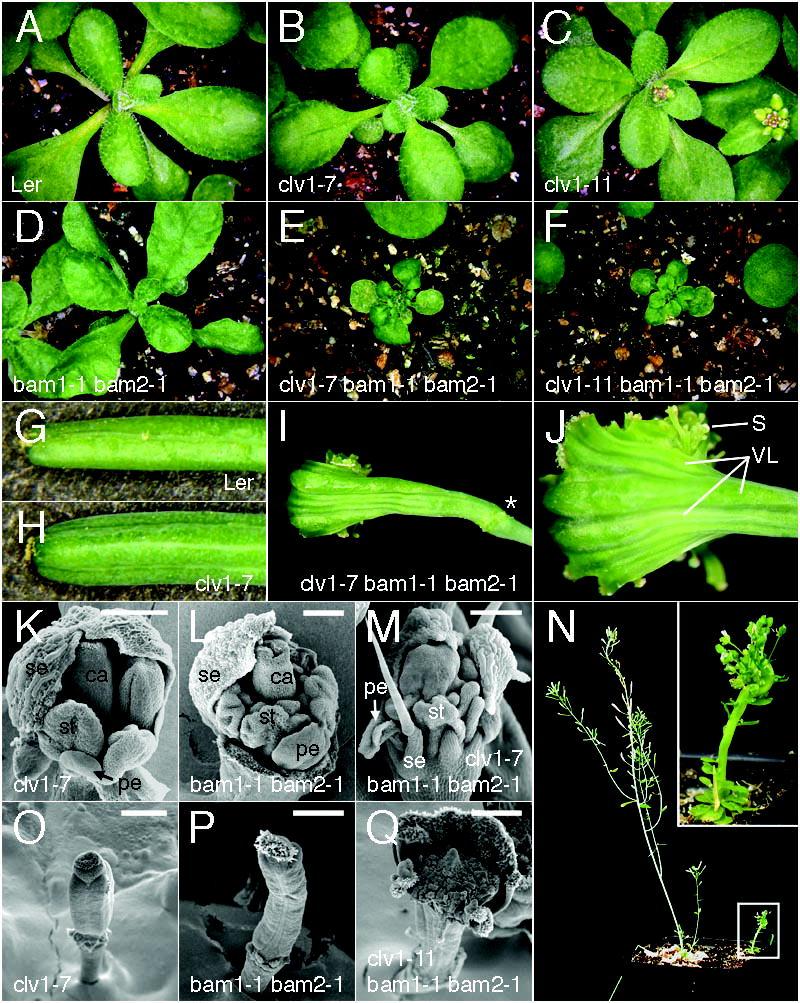
17 Consistent with this, the bam1 bam2 clv1-1 and the bam1 bam2 clv1-4 triple mutant plants were enhanced compared to the corresponding clv1 single mutant, and displayed phenotypes similar to bam1 bam2 clv1-null triple mutants (Table 2). Novel phenotypes in bam1 bam2 clv triple mutants: The shoot apical meristems of bam1 bam2 clv1 plants exhibited unique phenotypes not seen in any clv single mutant. These meristems occasionally lost organogenesis in portions of the meristem periphery (Figures 1M and 1N). Furthermore, bam1 bam2 clv1 shoot apical meristems, and to a lesser degree, clv1 bam1 shoot apical merisems, developed patches of differentiated cells (in one case, complete leaves) across the normally continuous field of undifferentiated meristem cells (Figures 1G, 1H, 1J, 1K, and 1L). Finally, bam1-1 bam2-1 clv1-11 and bam1-1 bam2-1 clv1-7 flowers developed an enormous indeterminate meristem in the center of each flower (Figure 2Q). In addition to the synergistic interactions observed in the shoot meristem, bam1-1 bam2-1 clv1-11 and bam1-1 bam2-1 clv1-7 triple mutants displayed a number of novel phenotypes outside of the meristem (Figure 2). bam1-1 bam2-1 clv1-11 and bam1-1 bam2-1 clv1-7 plants were much smaller than wild-type and bam1-1 bam2-1 plants, even when they were fully grown (Figures 2E, 2F, and 2N; data not shown). bam1-1 bam2-1 clv1-11 and bam1-1 bam2-1 clv1-7 plants had very small leaves and the number of rosette leaves was increased. The stems of the primary inflorescence of clv1-11 bam1-1 bam2-1 and clv1-7 bam1-1 17
18 bam2-1 plants were also extremely thick (Figure 2N; see below). The sepals, petals and stamens of bam1-1 bam2-1 clv1-11 and bam1-1 bam2-1 clv1-7 flowers were sometimes filamentous and generally looked less developed than those of bam1 bam2 flowers (Figure 2M; data not shown). The fruits of bam1-1 bam2-1 clv1-11 and bam1-1 bam2-1 clv1-7 plants were very short, however the pedicels were longer than that of wild type (Figure 2N). The replacement of valve tissue by replum tissue (valvelessness) has been previously observed in clv2 mutants (KAYES and CLARK 1998). This clv2 phenotype is enhanced by clv1 and clv3 mutants. Valvelessness was also strongly enhanced in the bam1 bam2 clv1 triple mutant, such that every bam1-1 bam2-1 clv1-11 and bam1-1 bam2-1 clv1-7 flower had missing or incomplete valves (Table 3). In addition, large sections of the gynoecia of these plants frequently lacked all valve character. We also observed large valveless regions that developed very thin stripes of valve-like tissue (Figures 2I and 2J). None of these synergistic shoot meristem or organ phenotypes were seen in either bam1 bam2 clv2 plants or bam1 bam2 clv3 plants (Table 3; Figure 1; data not shown), indicating a specific role for CLV1 outside the meristem. This represents a novel range of developmental pathways regulated by CLV1, and prompts consideration of broad functions for the orthologous rice FON1 and maize thick tassel dwarf1 (td1), both of whose expression appears broader than CLV1 (BOMMERT et al. 2005; SUZAKI et al. 2004). A defect in internal stem differentiation was observed in bam1 bam2 combinations with all clv mutations. clv mutations normally result in increased 18
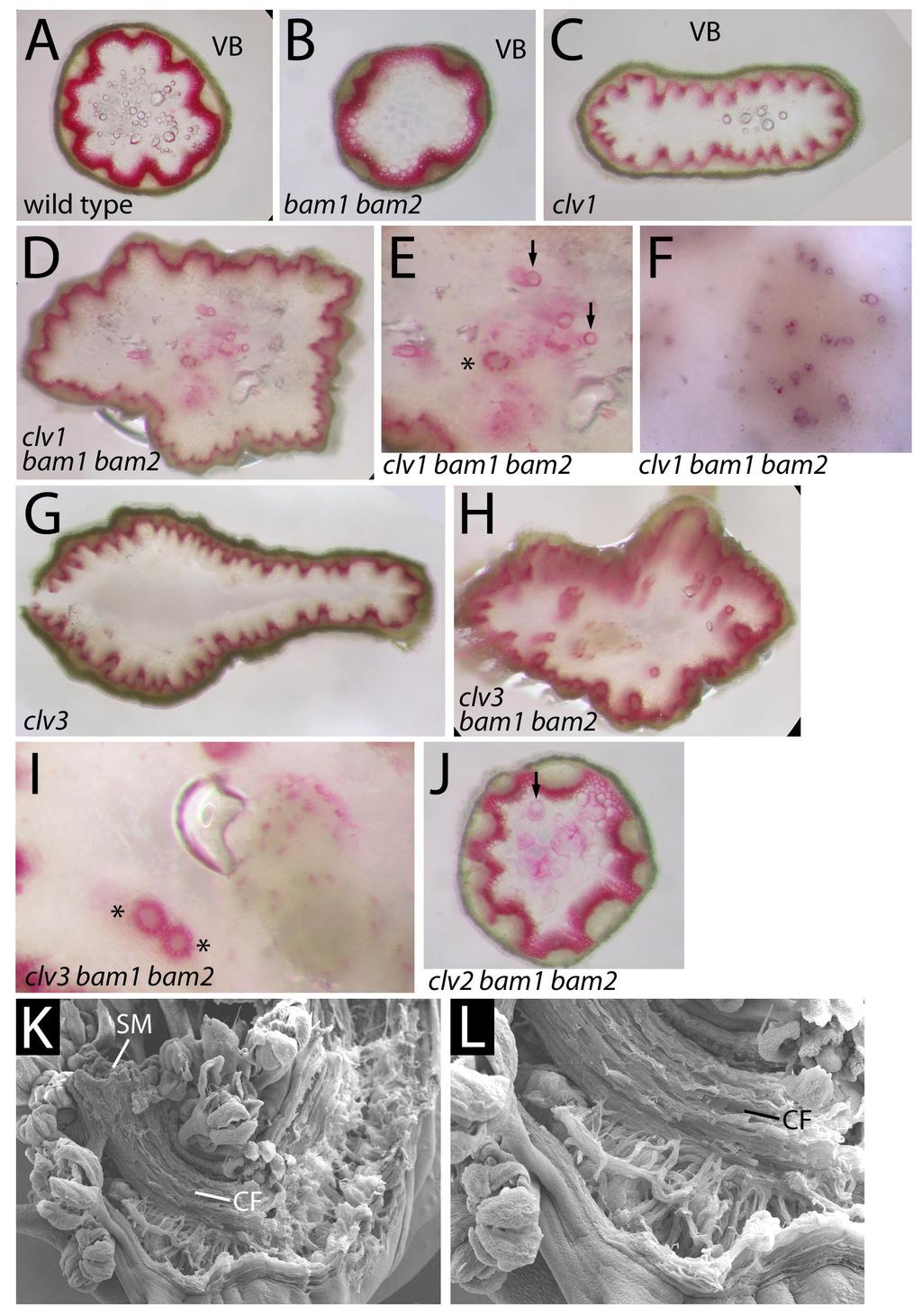
19 vascular bundle number, and rarely vascular bundles located away from the periphery of the stem, presumably a result of the enlarged shoot meristem (L. Doil and S. E. Clark, unpublished; Figure 3). In various bam1 bam2 clv mutant combinations, a more dramatic patterning defect was observed. Single, large ectopic lignified cells, as well as circular rings of smaller lignified cells were observed in the center of the stem (Figure 3D-F,H-J). Furthermore, in stronger clv mutant combinations, large portions of the internal stem were composed of poorly vacuolated cells with a mixture of lignified and non-lignified cells (Figure 3F,I). Stems of plants with this phenotype would occasionally break open during growth, revealing a fiber-like consistency to these abnormally developing stem regions (Figure 3K,L). What role defects in meristem development and what role defect in later stem differentiation play in this phenotype is unclear. 19
20 DISCUSSION A model for BAM function within the meristem: We investigated the genetic interactions of bam null alleles with various clv mutations. bam1 and bam2 exhibit dramatically different genetic interactions with clv1, clv2, and clv3, resulting in enhancement, no effect, and suppression, respectively. This indicates that the effects of BAM1 and BAM2 on stem cell homeostasis are integral to the CLV signaling pathway and do not represent a separate pathway regulating meristem development. bam1 bam2 mutations significantly suppressed the stem cell accumulation in clv3-2 flowers, consistent with a requirement for BAM receptors in stem cell maintenance. In contrast, clv2 was unaltered by bam1 bam2 within the meristem. Given previous data suggesting that CLV2 is required in part for CLV1 protein accumulation, we propose that BAM1 and BAM2 function within the meristem is dependent on CLV2. These results suggest that the clv2 phenotype, which is the strongest of any receptor null, would result from the combined reduction of CLV1 and BAM function. Unexpectedly, when combined with clv1 null alleles, both bam1 and bam2 strongly enhanced the Clv phenotype, and the triple mutant exhibited very severe defects in stem cell homeostasis. Thus bam mutants have an opposite effect on stem cell accumulation in a clv1 CLV3 background compared to a CLV1 CLV3 or a CLV1 clv3 background, raising the question of how these data can be reconciled. 20
21 A previous report has indicated that CLV1 likely acts to sequester the ligand CLV3 (LENHARD and LAUX 2003). One speculative possibility is that BAM receptors also act to sequester CLE polypeptide ligand(s) (Figure 4B-E). We have shown that many CLE-containing proteins can replace CLV3 function when expressed within the meristem, and some can act through CLV1 (NI and CLARK 2006). Furthermore, the CLE domain of these proteins appears to be proteolytically released as the active signaling molecule, and this domain is well conserved among Arabidopsis CLE-containing proteins (COCK and MCCORMICK 2001; DEYOUNG and CLARK 2001, Ito, 2006 #2620; FIERS et al. 2005; FIERS et al. 2004; KONDO et al. 2006). Indeed, multiple CLE domains can bind to the CLV1 extracellular domain (OGAWA et al. 2008). Over 30 CLE-containing proteins are expressed in large number of different tissues, including the meristem periphery, and evidence indicates that these peptides can diffuse over many cell diameters (FIERS et al. 2005; LENHARD and LAUX 2003; SHARMA et al. 2003). Thus, insulating the meristem center from CLE peptides diffusing from outside the meristem may be critical to maintain the proper feedback between WUS expression and CLV1 signaling (BRAND et al. 2000; SCHOOF et al. 2000). We suggest a model in which one of the functions of BAM receptors on the meristem periphery is to sequester CLE ligands and insulate the meristem center from extraneous signals (Figure 4B-E). In the absence of BAM receptors (i.e., in bam mutants), ectopic CLE signal would diffuse to the meristem center. This signal would not be subjected to the feedback WUS/CLV3 loop that normally regulates CLV3 expression and could lead to over-activation of CLV1 and 21
22 meristem reduction or termination. Furthermore, this ectopic CLE signal should be able to replace, in part, CLV3 function. Finally, the impact of this ectopic CLE signal resulting from bam mutations would only have an impact in a CLV1 background. Thus, the model explains the reduced meristem size of bam mutants, the bam suppression of clv3, and well as the requirement of CLV1 for meristem reduction in bam mutant backgrounds. While consistent with the genetic data, this model is quite speculative. We have no evidence of which CLE peptide might bind BAM receptors, and whether binding alone, as opposed to binding and signaling, is sufficient for this proposed BAM activity. One possible test as CLE expression data becomes available would be to inactivate candidate CLEs expressed on the meristem flanks in a bam mutant background. Another possible test would be to determine if CLE-binding, but non-functional BAM receptors expressed specifically on the meristem flanks could carry out BAM function there. These experiments are complicated by the exquisite sensitivity of the meristem center to BAM expression (see below), and the tendency of nonfunctional receptor-kinases to act as dominant-negative isoforms {Williams, 1997 #1555; Shpak, 2003 #2323; Diévart, 2003 #2328; Diévart, 2006 #2725} The enhancement of clv1 null alleles by bam mutations could indicate that while BAM receptors are at best expressed at low levels within the meristem center, they are redundant with CLV1 there (Figure 4A). clv1 dominant-negative alleles might be so effective in blocking signaling because there would theoretically be much more CLV1 than BAM receptor present in the meristem center. Redundant BAM/CLV1 function in the meristem center would explain the 22
23 observation that single bam mutations had no effect on the dominant-negative clv1-1 allele. Interestingly, the LRR missense allele clv1-4 did not display the same interaction with bam mutants, raising the possibility that its mechanism of dominant-negative action may differ from that of the kinase-domain missense mutants. CLV1/BAM synergy: Despite previous extensive phenotypic and expression analysis suggesting CLV1 function is meristem-specific, genetic analysis with bam mutants suggested otherwise. bam1 bam2 clv1 triple mutants exhibited many phenotypes not seen in any other genetic combination. These included strong dwarfism, filamentous floral organs, and loss of recognizable valve tissue. None of these phenotypes were seen in bam1 bam2 clv2 or bam1 bam2 clv3 plants, indicating that the phenotypes were not indirect consequences of defects in meristem development. This suggests that CLV1 plays a role in regulating the development of many organs in parallel with both BAM1 and BAM2. In addition, synergistic interactions were seen in the development of clv1 bam1 bam2 shoot meristems. Here, we observed differentiated cells, or even organs in the center of the meristem, and the occasional loss of organogenesis on the flanks of the meristem. These phenotypes are reminiscent of the effects of corona mutants and PLL1 over-expression, both of which enhance clv mutants, leading to a loss of organogenesis at the meristem periphery, massive stem cell accumulation, and predicted eventual loss of meristem identity resulting 23
24 in differentiation of the entire apex (GREEN et al. 2005; SONG et al. 2006). Alternatively, the loss of organogenesis might reflect a function for BAM1 and BAM2 on the periphery of the meristem, where the corresponding genes are expressed at relatively high levels. Determining the precise developmental roles for BAM1 and BAM2 in the meristem periphery as well as in the various plant organs is an essential next step in uncovering the function of these broadranging receptors. Genetic background effects: One potential complication of our analysis is that the bam1 alleles were isolated in the Columbia (Col) background, while all of the clv alleles were isolated or backcrossed into the Landsberg er (Ler) background (CLARK et al. 1993; CLARK et al. 1995; DEYOUNG et al. 2006; DIéVART et al. 2003; KOORNNEEF et al. 1983). We have shown previously that crosses between clv alleles in the Ler and Col backgrounds can have subtle, but measurable effects on the meristem phenotype (DIéVART et al. 2003). Two lines of evidence, however, indicate that background effects are not the source of the genetic interactions that we observed. First, the effects caused by genetic background are much smaller in magnitude than those observed here. Second, and most importantly, bam2-1 and bam2-3 alleles were isolated in the Ler background, and they exhibit the same type of interactions with each clv allele as observed for bam1. 24
25 ACKNOWLEDGMENTS This work was supported by a grant from the National Institutes of Health (R01GM62962) to S.E.C. B.J.D. was supported in part by the National Institutes of Health Cellular Biotechnology Training Program (5 T32 GM08353). 25
26 LITERATURE CITED BOMMERT, P., C. LUNDE, J. NARDMANN, E. VOLLBRECHT, M. RUNNING et al., 2005 thick tassel dwarf1 encodes a putative maize ortholog of the Arabidopsis CLAVATA1 leucine-rich repeat receptor-like kinase. Development 132: BRAND, U., J. C. FLETCHER, M. HOBE, E. M. MEYEROWITZ and R. SIMON, 2000 Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science 289: BYRNE, M. E., C. A. KIDNER and R. A. MARTIENSSEN, 2003 Plant stem cells: divergent pathways and common themes in shoots and roots. Curr Opin Genet Dev 13: CARLES, C. C., and J. C. FLETCHER, 2003 Shoot apical meristem maintenance: the art of a dynamic balance. Trends Plant Sci 8: CLARK, S. E., 1997 Organ formation at the vegetative shoot meristem. Plant Cell 9: CLARK, S. E., M. P. RUNNING and E. M. MEYEROWITZ, 1993 CLAVATA1, a regulator of meristem and flower development in Arabidopsis. Development 119: CLARK, S. E., M. P. RUNNING and E. M. MEYEROWITZ, 1995 CLAVATA3 is a specific regulator of shoot and floral meristem development affecting the same processes as CLAVATA1. Development 121:
27 CLARK, S. E., R. W. WILLIAMS and E. M. MEYEROWITZ, 1997 The CLAVATA1 gene encodes a putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell 89: COCK, J. M., and S. MCCORMICK, 2001 A large family of genes that share homology with CLAVATA3. Plant Physiol 126: DEYOUNG, B. J., K. L. BICKLE, K. J. SCHRAGE, P. MUSKETT, K. PATEL et al., 2006 The CLAVATA1-related BAM1, BAM2 and BAM3 receptor kinase-like proteins are required for meristem function in Arabidopsis. Plant J 45: DEYOUNG, B. J., and S. E. CLARK, 2001 Signaling through the CLAVATA1 receptor complex. Plant Mol Biol 46: DIéVART, A., M. DALAL, F. E. TAX, A. D. LACEY, A. HUTTLY et al., 2003 CLAVATA1 dominant-negative alleles reveal functional overlap between multiple receptor kinases that regulate meristem and organ development. Plant Cell 15: FIERS, M., E. GOLEMIEC, J. XU, L. VAN DER GEEST, R. HEIDSTRA et al., 2005 The 14- amino acid CLV3, CLE19, and CLE40 peptides trigger consumption of the root meristem in Arabidopsis through a CLAVATA2-dependent pathway. Plant Cell 17: FIERS, M., G. HAUSE, K. BOUTILIER, E. CASAMITJANA-MARTINEZ, D. WEIJERS et al., 2004 Mis-expression of the CLV3/ESR-like gene CLE19 in Arabidopsis leads to a consumption of root meristem. Gene 327:
28 GREEN, K. A., M. J. PRIGGE, R. B. KATZMAN and S. E. CLARK, 2005 CORONA, a Member of the Class III Homeodomain-Leucine Zipper Gene Family in Arabidopsis, Regulates Stem Cell Specification and Organogenesis. Plant Cell In Press. GROSS-HARDT, R., and T. LAUX, 2003 Stem cell regulation in the shoot meristem. J Cell Sci 116: HORD, C. L., C. CHEN, B. J. DEYOUNG, S. E. CLARK and H. MA, 2006 The BAM1/BAM2 receptor-like kinases are important regulators of Arabidopsis early anther development. Plant Cell 18: ITO, Y., I. NAKANOMYO, H. MOTOSE, K. IWAMOTO, S. SAWA et al., 2006 Dodeca-CLE peptides as suppressors of plant stem cell differentiation. Science 313: JEONG, S., A. E. TROTOCHAUD and S. E. CLARK, 1999 The Arabidopsis CLAVATA2 gene encodes a receptor-like protein required for the stability of the CLAVATA1 receptor-like kinase. Plant Cell 11: KAYES, J. M., and S. E. CLARK, 1998 CLAVATA2, a regulator of meristem and organ development in Arabidopsis. Development 125: KONDO, T., S. SAWA, A. KINOSHITA, S. MIZUNO, T. KAKIMOTO et al., 2006 A plant peptide encoded by CLV3 identified by in situ MALDI-TOF MS analysis. Science 313: KOORNNEEF, M., J. VAN EDEN, C. J. HANHART, P. STAM, F. J. BRAAKSMA et al., 1983 Linkage map of Arabidopsis thaliana. J. Hered. 74:
29 LAUX, T., K. F. MAYER, J. BERGER and G. JURGENS, 1996 The WUSCHEL gene is required for shoot and floral meristem integrity in Arabidopsis. Development 122: LENHARD, M., and T. LAUX, 2003 Stem cell homeostasis in the Arabidopsis shoot meristem is regulated by intercellular movement of CLAVATA3 and its sequestration by CLAVATA1. Development 130: MAYER, K. F., H. SCHOOF, A. HAECKER, M. LENHARD, G. JURGENS et al., 1998 Role of WUSCHEL in regulating stem cell fate in the Arabidopsis shoot meristem. Cell 95: NI, J., and S. E. CLARK, 2006 Evidence for functional conservation, sufficiency, and proteolytic processing of the CLAVATA3 CLE domain. Plant Physiol 140: OGAWA, M., H. SHINOHARA, Y. SAKAGAMI and Y. MATSUBAYASHI, 2008 Arabidopsis CLV3 peptide directly binds CLV1 ectodomain. Science 319: 294. PRIGGE, M. J., D. OTSUGA, J. M. ALONSO, J. R. ECKER, G. N. DREWS et al., 2005 Class III Homeodomain-Leucine Zipper Gene Family Members Have Overlapping, Antagonistic, and Distinct Roles in Arabidopsis Development. Plant Cell 17: SCHOOF, H., M. LENHARD, A. HAECKER, K. F. MAYER, G. JURGENS et al., 2000 The stem cell population of Arabidopsis shoot meristems in maintained by a regulatory loop between the CLAVATA and WUSCHEL genes. Cell 100:
30 SHARMA, V. K., J. RAMIREZ and J. C. FLETCHER, 2003 The Arabidopsis CLV3-like (CLE) genes are expressed in diverse tissues and encode secreted proteins. Plant Mol Biol 51: SONG, S.-K., and S. E. CLARK, 2005 POL and related phosphatases are dosagesensitive regulators of meristem and organ development in Arabidopsis. Dev Biol in press. SONG, S. K., M. M. LEE and S. E. CLARK, 2006 POL and PLL1 phosphatases are CLAVATA1 signaling intermediates required for Arabidopsis shoot and floral stem cells. Development 133: SUZAKI, T., M. SATO, M. ASHIKARI, M. MIYOSHI, Y. NAGATO et al., 2004 The gene FLORAL ORGAN NUMBER1 regulates floral meristem size in rice and encodes a leucine-rich repeat receptor kinase orthologous to Arabidopsis CLAVATA1. Development 131: TROTOCHAUD, A. E., T. HAO, W. GUANG, Z. YANG and S. E. CLARK, 1999 The CLAVATA1 receptor-like kinase requires CLAVATA3 for its assembly into a signaling complex that includes KAPP and a Rho-related protein. Plant Cell 11: YU, L. P., A. K. MILLER and S. E. CLARK, 2003 POLTERGEIST Encodes a Protein Phosphatase 2C that Regulates CLAVATA Pathways Controlling Stem Cell Identity at Arabidopsis Shoot and Flower Meristems. Curr Biol 13:
31 YU, L. P., E. J. SIMON, A. E. TROTOCHAUD and S. E. CLARK, 2000 POLTERGEIST functions to regulate meristem development downstream of the CLAVATA loci. Development 127:
32 TABLES TABLE 1. Phenotypic interactions of bam1 and bam2 mutations with clv3 and clv2 mutations. Mean Genotype carpels/flower SE a n b p c Ler d bam2-1 d bam2-3 d bam1-1 d bam1-3 d bam1-1 bam2-1 d bam1-3 bam2-3 d clv clv3-2 bam <0.001 clv3-2 bam <0.001 clv3-2 bam1-1 bam <0.001 clv clv2-1 bam <0.001 clv2-1 bam <0.001 clv2-1 bam1-1 bam a Standard error. 32
33 b Number of flowers evaluated. c p value calculated using Student s t-test comparing clv bam1, clv bam2, or clv bam1 bam2 to the respective clv single mutant. d Data from DeYoung et al. (2006). 33
34 TABLE 2. Phenotypic interactions of bam1 and bam2 mutations with null and dominant-negative clv1 alleles. Mean Genotype carpels/flower SE a n b p c clv clv1-7 bam <0.001 clv1-7 bam <0.001 clv1-7 bam <0.001 clv1-7 bam <0.001 clv1-7 bam1-1 bam <0.001 clv clv1-11 bam <0.001 clv1-11 bam <0.001 clv1-11 bam <0.001 clv1-11 bam <0.001 clv1-11 bam1-1 bam <0.001 clv clv1-1 bam clv1-1 bam clv1-1 bam1-1 bam <0.001 clv
35 clv1-4 bam <0.001 clv1-4 bam <0.001 clv1-4 bam1-1 bam <0.001 a Standard error. b Number of flowers evaluated. c p value calculated using Student s t-test comparing clv1 bam1, clv1 bam2, or clv1 bam1 bam2 to the respective clv1 single mutant. 35
36 TABLE 3. bam1 and bam2 affect valvelessness Genotype % valveless a n b Ler clv clv1-7 bam clv1-7 bam clv1-7 bam1-1 bam clv clv2-1 bam clv2-1 bam clv2-1 bam1-1 bam a Percent of flowers with at least one missing or incomplete valve. Missing valves were defined as the replacement of a complete valve in the gynoecium by replum tissue. Incomplete valves were defined as valves that occupied less than 75% of the distance from the receptacle to the stigma. b Number of flowers evaluated. 36
37 FIGURE LEGENDS FIGURE 1. bam1 and bam2 enhance meristem enlargement by clv1 and function synergistically with clv1 resulting in novel phenotypes. Scanning electron micrographs of Ler (A), clv1-11 (B), clv1-11 bam2-3 (C), clv1-11 bam1-3 (D), clv1-7 (E), clv1-7 bam2-3 (F), clv1-7 bam1-3 (G, H), clv1-11 bam1-1 bam2-1 (I, J), clv1-7 bam1-1 bam2-1 (K-N) Older flowers were removed for clarity. Panels H, L, and N correspond to the boxed regions in G, K, and M, respectively. Differentiated cells surrounded by undifferentiated merstiem cells are indicated with an arrow (H and L). Lack of flower primordia are indicated by an arrow (N). A, B, E, and F are shown at the same magnification (scale bar 50μm). Scale bars, 50μm (H and L); 100μm (C, D, and N); 250μm (J,); 500μM (G, I, K, and M). FIGURE 2. clv1 bam1 bam2 plants exhibit novel phenotypes in floral meristem and non-meristem tissues. Rosettes of Ler (A), clv1-7 (B), clv1-11 (C), bam1-1 bam2-1 (D), clv1-7 bam1-1 bam2-1 (E), and clv1-11 bam1-1 bam2-1 plants 14 days after germination. Gynoecia of Ler (G), clv1-7 (H), and clv1-7 bam1-1 bam2-1 (I,J) plants. Scanning electron micrographs of clv1-7 (K), bam1-1 bam2-1 (L), and clv1-7 bam1-1 bam2-1 (M) young flowers are shown. Some sepals were removed from the clv1-7 (K) and bam1-1 bam2-1 (L) flower to reveal the inner organs. No organs were removed from the clv1-7 bam1-1 bam2-1 flower (M). clv1-7 bam1-1 37
38 bam2-1 plant (right, boxed) beside a sibling plant (left) that is either heterozygous or homozygous wild-type at both BAM1 and BAM2 loci (N). Close-up of clv1-7 bam1-1 bam2-1 plant (N, inset). Scanning electron micrographs of clv1-7 (O), bam1-1 bam1-2 (P), and clv1-11 bam1-1 bam1-2 gynoecia are shown. A-F are shown at the same magnification as are G, H, and I. J is a close-up view of a clv1-7 bam1-1 bam2-1 gynoecium. Scalebars, 100 μm (L,M); 250μm (K); 500μm (O-Q). *, attachment site of sepals, petals and stamens; S stigmatic tissue; VL valve-like strips; se, sepal primordium; pe, petal primordium; st, stamen primordium; ca, carpel primordium. FIGURE 3. Synergistic vascular patterning defects clv bam1 bam2 mutants. Cross sections of wild type Ler (A), bam1-1 bam2-1 (B), clv1-1 (C), clv1-1 bam1-1 bam2-1 (D-F), clv3-2 (G), clv3-2 bam1-1 bam2-1 (H), clv2-1 (I), clv2-1 bam1-1 bam2-1 (J) stems stained with phloroglucinol (red) to detect lignified tissues. Scanning electron micrographs of clv1-11 bam1-1 bam2-1 mutants (K, L) show stems that split open during growth to reveal a fiber-like inner structure. Portions of the stem were removed to better visualize this structure. VB, vascular bundle, SM, shoot meristem. Arrows indicate large lignified cells, asterisks indicate rings of smaller lignified cells. 38
39 FIGURE 4. A model for BAM function through signal transduction and ligand sequestration. (A) In wild-type cell layer three (L3) meristem cells, CLV1 is expressed at higher levels than BAM proteins and is the primary transducer of CLE signal. CLV1/CLV1 and CLV1/BAM complexes are consistent with the dominantnegative function of many clv1 alleles and their interaction with bam mutations. In clv1-1 L3 cells, BAM signaling may be blocked by interference of the mutant clv1-1 kinase domain. Residual signaling may come from a partially active BAM/clv1-1 heterodimer and/or the presumably small amount of BAM/BAM homodimers (not shown). In the absence of CLV1 (clv1-null), BAM proteins are available to transduce CLE signal, albeit at a reduced level compared to WT cells. To simplify the diagram, CLV2 and other components have not been included. (B) In a wild-type shoot meristem, CLV3 signals primarily through CLV1, but also through BAM1 and BAM2 to regulate the WUS expression domain. BAM1 and BAM2 also function on the flanks of the meristem where they sequester CLV3-related CLE ligands, creating a buffer around the meristem center. (C) In a clv3 apex, WUS repression is lost, leading to an expansion of the WUS-expression domain and an enlargement of the meristem, while CLE ligands outside of the meristem are sequestered. (D) In a bam1 bam2 apex, CLE ligands are no longer sequestered, leading to over-activation of CLV1 and reduction in the meristem size, even meristem termination in bam1 bam2 bam3 triple mutants. (E) In a clv3 bam1 bam2 apex, loss of CLV3 is partially balanced through activation of CLV1 by unsequestered CLE ligands resulting in partial 39
40 WUS expression control. (F) A model depicting genetic pathways through which CLV1, BAM1, and BAM2 function. 40

41
42
43
44
Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family
, HAESA, ERECTA-family") Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family GENES & DEVELOPMENT (2000) 14: 108 117 INTRODUCTION Flower Diagram INTRODUCTION Abscission In plant, the process by which a plant
Leucine-rich repeat receptor-like kinases (LRR-RLKs), HAESA, ERECTA-family GENES & DEVELOPMENT (2000) 14: 108 117 INTRODUCTION Flower Diagram INTRODUCTION Abscission In plant, the process by which a plant
CLE peptide ligands ; plant polypeptide signaling molecules
 CLE peptide ligands ; plant polypeptide signaling molecules ligand receptor-like kinase signalling pathways ; such signalling cascades for plant growth and development Model for CLV3 peptide signaling
CLE peptide ligands ; plant polypeptide signaling molecules ligand receptor-like kinase signalling pathways ; such signalling cascades for plant growth and development Model for CLV3 peptide signaling
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature12791 Supplementary Figure 1 (1/3) WWW.NATURE.COM/NATURE 1 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Figure 1 (2/3) 2 WWW.NATURE.COM/NATURE SUPPLEMENTARY
SUPPLEMENTARY INFORMATION doi:10.1038/nature12791 Supplementary Figure 1 (1/3) WWW.NATURE.COM/NATURE 1 RESEARCH SUPPLEMENTARY INFORMATION Supplementary Figure 1 (2/3) 2 WWW.NATURE.COM/NATURE SUPPLEMENTARY
The Plant Cell, November. 2017, American Society of Plant Biologists. All rights reserved
 The Genetics of Floral Development Teaching Guide Overview The development of flowers in angiosperm plants provided a critical evolutionary advantage, allowing more options for pollen dispersal and seed
The Genetics of Floral Development Teaching Guide Overview The development of flowers in angiosperm plants provided a critical evolutionary advantage, allowing more options for pollen dispersal and seed
Identification of Components Controlling Meristem Homeostasis Lindsey Ann Gish
 Identification of Components Controlling Meristem Homeostasis by Lindsey Ann Gish A dissertation submitted in the partial fulfillment of the requirements for the degree of Doctor of Philosophy (Molecular,
Identification of Components Controlling Meristem Homeostasis by Lindsey Ann Gish A dissertation submitted in the partial fulfillment of the requirements for the degree of Doctor of Philosophy (Molecular,
CLE peptide ligands and their roles in establishing meristems Martijn Fiers 1, Ka Lei Ku 2 and Chun-Ming Liu 2
 CLE peptide ligands and their roles in establishing meristems Martijn Fiers 1, Ka Lei Ku 2 and Chun-Ming Liu 2 Research in the past decade revealed that peptide ligands, also called peptide hormones, play
CLE peptide ligands and their roles in establishing meristems Martijn Fiers 1, Ka Lei Ku 2 and Chun-Ming Liu 2 Research in the past decade revealed that peptide ligands, also called peptide hormones, play
Chapter 18 Lecture. Concepts of Genetics. Tenth Edition. Developmental Genetics
 Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
Chapter 18 Lecture Concepts of Genetics Tenth Edition Developmental Genetics Chapter Contents 18.1 Differentiated States Develop from Coordinated Programs of Gene Expression 18.2 Evolutionary Conservation
The mode of development in animals and plants is different
 The mode of development in animals and plants is different Outcome of animal embryogenesis is a mini edition of the adult Outcome of plant embryogenesis is a simple structure with -root apical meristem
The mode of development in animals and plants is different Outcome of animal embryogenesis is a mini edition of the adult Outcome of plant embryogenesis is a simple structure with -root apical meristem
Receptor Kinases in Plant Meristem Development
 Receptor Kinases in Plant Meristem Development Yvonne Stahl and R udiger Simon Abstract Meristems are groups of cells that promote shoot and root growth, including the growth of new organs, and control
Receptor Kinases in Plant Meristem Development Yvonne Stahl and R udiger Simon Abstract Meristems are groups of cells that promote shoot and root growth, including the growth of new organs, and control
Plants ranging from the small weed Arabidopsis to the giant
 Multiple feedback loops through cytokinin signaling control stem cell number within the Arabidopsis shoot meristem Sean P. Gordon, Vijay S. Chickarmane, Carolyn Ohno 1, and Elliot M. Meyerowitz 2 Division
Multiple feedback loops through cytokinin signaling control stem cell number within the Arabidopsis shoot meristem Sean P. Gordon, Vijay S. Chickarmane, Carolyn Ohno 1, and Elliot M. Meyerowitz 2 Division
Biological Roles of Cytokinins
 Direct Control of Shoot Meristem Activity by a Cytokinin-Activating Enzyme By Kurakawa et. Al. Published in Nature Presented by Boyana Grigorova Biological Roles of Cytokinins Cytokinins are positive regulators
Direct Control of Shoot Meristem Activity by a Cytokinin-Activating Enzyme By Kurakawa et. Al. Published in Nature Presented by Boyana Grigorova Biological Roles of Cytokinins Cytokinins are positive regulators
Signaling In and Out: Control of Cell Division and Differentiation in the Shoot and Root
 The Plant Cell, S265 S276, Supplement 2002, www.plantcell.org 2002 American Society of Plant Biologists Signaling In and Out: Control of Cell Division and Differentiation in the Shoot and Root Keiji Nakajima
The Plant Cell, S265 S276, Supplement 2002, www.plantcell.org 2002 American Society of Plant Biologists Signaling In and Out: Control of Cell Division and Differentiation in the Shoot and Root Keiji Nakajima
Simon Scofield, Walter Dewitte, and James AH Murray* School of Biosciences; Cardiff University; Cardiff, UK
 Short Communication Plant Signaling & Behavior 9, e28934; April; 2014 Landes Bioscience Short Communication STM sustains stem cell function in the Arabidopsis shoot apical meristem and controls KNOX gene
Short Communication Plant Signaling & Behavior 9, e28934; April; 2014 Landes Bioscience Short Communication STM sustains stem cell function in the Arabidopsis shoot apical meristem and controls KNOX gene
A MicroRNA as a Translational Repressor of APETALA2 in Arabidopsis Flower Development
 A MicroRNA as a Translational Repressor of APETALA2 in Arabidopsis Flower Development Xuemei Chen Waksman Institute, Rutgers University, Piscataway, NJ 08854, USA. E-mail: xuemei@waksman.rutgers.edu Plant
A MicroRNA as a Translational Repressor of APETALA2 in Arabidopsis Flower Development Xuemei Chen Waksman Institute, Rutgers University, Piscataway, NJ 08854, USA. E-mail: xuemei@waksman.rutgers.edu Plant
Outline. Leaf Development. Leaf Structure - Morphology. Leaf Structure - Morphology
 Outline 1. Leaf Structure: Morphology & Anatomy 2. Leaf Development A. Anatomy B. Sector analysis C. Leaf Development Leaf Structure - Morphology Leaf Structure - Morphology 1 Leaf Structure - Morphology
Outline 1. Leaf Structure: Morphology & Anatomy 2. Leaf Development A. Anatomy B. Sector analysis C. Leaf Development Leaf Structure - Morphology Leaf Structure - Morphology 1 Leaf Structure - Morphology
BILD7: Problem Set. 2. What did Chargaff discover and why was this important?
 BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
BILD7: Problem Set 1. What is the general structure of DNA? 2. What did Chargaff discover and why was this important? 3. What was the major contribution of Rosalind Franklin? 4. How did solving the structure
Regulation of Vascular Development by CLE Peptide-receptor Systems
 Journal of Integrative Plant Biology 2010, 52 (1): 8 16 Invited Expert Review Regulation of Vascular Development by CLE Peptide-receptor Systems Yuki Hirakawa, Yuki Kondo and Hiroo Fukuda Department of
Journal of Integrative Plant Biology 2010, 52 (1): 8 16 Invited Expert Review Regulation of Vascular Development by CLE Peptide-receptor Systems Yuki Hirakawa, Yuki Kondo and Hiroo Fukuda Department of
Mitogen-Activated Protein Kinase Regulated by the CLAVATA Receptors Contributes to Shoot Apical Meristem Homeostasis
 Special Issue Regular Paper Mitogen-Activated Protein Kinase Regulated by the CLAVATA Receptors Contributes to Shoot Apical Meristem Homeostasis Shigeyuki Betsuyaku 1, 2, 5, *, Fuminori Takahashi 3, 4,
Special Issue Regular Paper Mitogen-Activated Protein Kinase Regulated by the CLAVATA Receptors Contributes to Shoot Apical Meristem Homeostasis Shigeyuki Betsuyaku 1, 2, 5, *, Fuminori Takahashi 3, 4,
Nature Genetics: doi: /ng Supplementary Figure 1. The phenotypes of PI , BR121, and Harosoy under short-day conditions.
 Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Supplementary Figure 1 The phenotypes of PI 159925, BR121, and Harosoy under short-day conditions. (a) Plant height. (b) Number of branches. (c) Average internode length. (d) Number of nodes. (e) Pods
Nature Genetics: doi: /ng Supplementary Figure 1. The FIN and FAB genes act separately from the meristem maturation pathway.
 Supplementary Figure 1 The FIN and FAB genes act separately from the meristem maturation pathway. (a) Representative inflorescence from the compound inflorescence (s, defective in the homolog of Arabidopsis
Supplementary Figure 1 The FIN and FAB genes act separately from the meristem maturation pathway. (a) Representative inflorescence from the compound inflorescence (s, defective in the homolog of Arabidopsis
Curriculum vitae Xigang Liu
 Curriculum vitae Xigang Liu 1, EDUCATION: 09/1993-07/1997 B.S. Major: Biology. College of Life Sciences, Hebei Normal University Academic Degree Paper: RAPD analysis of Taigu genic male-sterile wheat and
Curriculum vitae Xigang Liu 1, EDUCATION: 09/1993-07/1997 B.S. Major: Biology. College of Life Sciences, Hebei Normal University Academic Degree Paper: RAPD analysis of Taigu genic male-sterile wheat and
Supplemental Data. Wang et al. (2014). Plant Cell /tpc
. Plant Cell /tpc") Supplemental Figure1: Mock and NPA-treated tomato plants. (A) NPA treated tomato (cv. Moneymaker) developed a pin-like inflorescence (arrowhead). (B) Comparison of first and second leaves from mock and
Supplemental Figure1: Mock and NPA-treated tomato plants. (A) NPA treated tomato (cv. Moneymaker) developed a pin-like inflorescence (arrowhead). (B) Comparison of first and second leaves from mock and
Peptide signalling in plant development:
 Peptide signalling in plant development: Functional analysis of CLE ligands in Arabidopsis Martijn Fiers Promotor: Prof. Dr. W. J. Stiekema Hoogleraar Genoom Informatica, Laboratorium voor Bioinformatica,
Peptide signalling in plant development: Functional analysis of CLE ligands in Arabidopsis Martijn Fiers Promotor: Prof. Dr. W. J. Stiekema Hoogleraar Genoom Informatica, Laboratorium voor Bioinformatica,
Nature Genetics: doi: /ng Supplementary Figure 1. ssp mutant phenotypes in a functional SP background.
 Supplementary Figure 1 ssp mutant phenotypes in a functional SP background. (a,b) Statistical comparisons of primary and sympodial shoot flowering times as determined by mean values for leaf number on
Supplementary Figure 1 ssp mutant phenotypes in a functional SP background. (a,b) Statistical comparisons of primary and sympodial shoot flowering times as determined by mean values for leaf number on
The BAM1/BAM2 Receptor-Like Kinases Are Important Regulators of Arabidopsis Early Anther Development W
 The Plant Cell, Vol. 18, 1667 1680, July 2006, www.plantcell.org ª 2006 American Society of Plant Biologists The BAM1/BAM2 Receptor-Like Kinases Are Important Regulators of Arabidopsis Early Anther Development
The Plant Cell, Vol. 18, 1667 1680, July 2006, www.plantcell.org ª 2006 American Society of Plant Biologists The BAM1/BAM2 Receptor-Like Kinases Are Important Regulators of Arabidopsis Early Anther Development
Regulation of Phosphate Homeostasis by microrna in Plants
 Regulation of Phosphate Homeostasis by microrna in Plants Tzyy-Jen Chiou 1 *, Kyaw Aung 1,2, Shu-I Lin 1,3, Chia-Chune Wu 1, Su-Fen Chiang 1, and Chun-Lin Su 1 Abstract Upon phosphate (Pi) starvation,
Regulation of Phosphate Homeostasis by microrna in Plants Tzyy-Jen Chiou 1 *, Kyaw Aung 1,2, Shu-I Lin 1,3, Chia-Chune Wu 1, Su-Fen Chiang 1, and Chun-Lin Su 1 Abstract Upon phosphate (Pi) starvation,
Plant Molecular and Cellular Biology Lecture 10: Plant Cell Cycle Gary Peter
 Plant Molecular and Cellular Biology Lecture 10: Plant Cell Cycle Gary Peter 9/10/2008 1 Learning Objectives Explain similarities and differences between fungal, mammalian and plant cell cycles Explain
Plant Molecular and Cellular Biology Lecture 10: Plant Cell Cycle Gary Peter 9/10/2008 1 Learning Objectives Explain similarities and differences between fungal, mammalian and plant cell cycles Explain
The Coch gene controls the subsequent differentiation of pea axial meristems into lateral structures
 The Coch gene controls the subsequent differentiation of pea axial meristems into lateral structures Rozov, S.M. 1, Institute of Cytology and Genetics SD RAS, Novosibirsk, Russia Voroshilova, V.A. 2, 2
The Coch gene controls the subsequent differentiation of pea axial meristems into lateral structures Rozov, S.M. 1, Institute of Cytology and Genetics SD RAS, Novosibirsk, Russia Voroshilova, V.A. 2, 2
Plant Structure, Growth, and Development
 Plant Structure, Growth, and Development Plant hierarchy: Cells Tissue: group of similar cells with similar function: Dermal, Ground, Vascular Organs: multiple kinds of tissue, very diverse function Organ
Plant Structure, Growth, and Development Plant hierarchy: Cells Tissue: group of similar cells with similar function: Dermal, Ground, Vascular Organs: multiple kinds of tissue, very diverse function Organ
Epigenetics and Flowering Any potentially stable and heritable change in gene expression that occurs without a change in DNA sequence
 Epigenetics and Flowering Any potentially stable and heritable change in gene expression that occurs without a change in DNA sequence www.plantcell.org/cgi/doi/10.1105/tpc.110.tt0110 Epigenetics Usually
Epigenetics and Flowering Any potentially stable and heritable change in gene expression that occurs without a change in DNA sequence www.plantcell.org/cgi/doi/10.1105/tpc.110.tt0110 Epigenetics Usually
Genetic interactions of the Arabidopsis flowering time gene FCA, with genes regulating floral initiation
 The Plant Journal (1999) 17(3), 231 239 Genetic interactions of the Arabidopsis flowering time gene FCA, with genes regulating floral initiation Tania Page 1,, Richard Macknight 1,, Chang-Hsien Yang 2
The Plant Journal (1999) 17(3), 231 239 Genetic interactions of the Arabidopsis flowering time gene FCA, with genes regulating floral initiation Tania Page 1,, Richard Macknight 1,, Chang-Hsien Yang 2
Cytokinin. Fig Cytokinin needed for growth of shoot apical meristem. F Cytokinin stimulates chloroplast development in the dark
 Cytokinin Abundant in young, dividing cells Shoot apical meristem Root apical meristem Synthesized in root tip, developing embryos, young leaves, fruits Transported passively via xylem into shoots from
Cytokinin Abundant in young, dividing cells Shoot apical meristem Root apical meristem Synthesized in root tip, developing embryos, young leaves, fruits Transported passively via xylem into shoots from
Bypass and interaction suppressors; pathway analysis
 Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
Bypass and interaction suppressors; pathway analysis The isolation of extragenic suppressors is a powerful tool for identifying genes that encode proteins that function in the same process as a gene of
GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL
 GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL Kelsey Hoth 1 Dr. Maria Ivanchenko 2 Bioresourse Research 1, Department of Botany and Plant Physiology 2, Oregon State University, Corvallis,
GENETIC ANALYSES OF ROOT SYSTEM DEVELOPMENT IN THE TOMATO CROP MODEL Kelsey Hoth 1 Dr. Maria Ivanchenko 2 Bioresourse Research 1, Department of Botany and Plant Physiology 2, Oregon State University, Corvallis,
The Function of the CLE Peptides in Plant Development and Plant- Microbe Interactions
 The Function of the CLE Peptides in Plant Development and Plant- Microbe Interactions Authors: Shigeyuki Betsuyaku, Shinichiro Sawa, and Masashi Yamada Source: The Arabidopsis Book, 2011(9) Published By:
The Function of the CLE Peptides in Plant Development and Plant- Microbe Interactions Authors: Shigeyuki Betsuyaku, Shinichiro Sawa, and Masashi Yamada Source: The Arabidopsis Book, 2011(9) Published By:
Analysis of Transgenic Tobacco with Overexpression of Arabidopsis WUSCHEL Gene
 Acta Botanica Sinica 2004, 46 (2): 224 229 http://www.chineseplantscience.com Analysis of Transgenic Tobacco with Overexpression of Arabidopsis WUSCHEL Gene LI Jun-Hua 1, 2, XU Yun-Yuan 1, CHONG Kang 1*,
Acta Botanica Sinica 2004, 46 (2): 224 229 http://www.chineseplantscience.com Analysis of Transgenic Tobacco with Overexpression of Arabidopsis WUSCHEL Gene LI Jun-Hua 1, 2, XU Yun-Yuan 1, CHONG Kang 1*,
Genetic Characterization and Functional Analysis of the GID1 Gibberellin Receptors in Arabidopsis W
 The Plant Cell, Vol. 18, 3399 3414, December 2006, www.plantcell.org ª 2006 American Society of Plant Biologists Genetic Characterization and Functional Analysis of the GID1 Gibberellin Receptors in Arabidopsis
The Plant Cell, Vol. 18, 3399 3414, December 2006, www.plantcell.org ª 2006 American Society of Plant Biologists Genetic Characterization and Functional Analysis of the GID1 Gibberellin Receptors in Arabidopsis
purpose of this Chapter is to highlight some problems that will likely provide new
 119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
119 Chapter 6 Future Directions Besides our contributions discussed in previous chapters to the problem of developmental pattern formation, this work has also brought new questions that remain unanswered.
GFP GAL bp 3964 bp
 Supplemental Data. Møller et al. (2009) Shoot Na + exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na + transport in Arabidopsis Supplemental Figure 1. Salt-sensitive
Supplemental Data. Møller et al. (2009) Shoot Na + exclusion and increased salinity tolerance engineered by cell type-specific alteration of Na + transport in Arabidopsis Supplemental Figure 1. Salt-sensitive
Supplementary Figure 1. Phenotype of the HI strain.
 Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Supplementary Figure 1. Phenotype of the HI strain. (A) Phenotype of the HI and wild type plant after flowering (~1month). Wild type plant is tall with well elongated inflorescence. All four HI plants
Arabidopsis thaliana. Lucia Strader. Assistant Professor, Biology
 Arabidopsis thaliana Lucia Strader Assistant Professor, Biology Arabidopsis as a genetic model Easy to grow Small genome Short life cycle Self fertile Produces many progeny Easily transformed HIV E. coli
Arabidopsis thaliana Lucia Strader Assistant Professor, Biology Arabidopsis as a genetic model Easy to grow Small genome Short life cycle Self fertile Produces many progeny Easily transformed HIV E. coli
WOX4 and WOX14 act downstream of the PXY receptor kinase to regulate plant vascular proliferation independently of any role in vascular organisation
 2224 Development 140, 2224-2234 (2013) doi:10.1242/dev.091314 2013. Published by The Company of Biologists Ltd WOX4 and WOX14 act downstream of the PXY receptor kinase to regulate plant vascular proliferation
2224 Development 140, 2224-2234 (2013) doi:10.1242/dev.091314 2013. Published by The Company of Biologists Ltd WOX4 and WOX14 act downstream of the PXY receptor kinase to regulate plant vascular proliferation
Plant Growth and Development
 Plant Growth and Development Concept 26.1 Plants Develop in Response to the Environment Factors involved in regulating plant growth and development: 1. Environmental cues (e.g., day length) 2. Receptors
Plant Growth and Development Concept 26.1 Plants Develop in Response to the Environment Factors involved in regulating plant growth and development: 1. Environmental cues (e.g., day length) 2. Receptors
Reading: Chapter 5, pp ; Reference chapter D, pp Problem set F
 Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Mosaic Analysis Reading: Chapter 5, pp140-141; Reference chapter D, pp820-823 Problem set F Twin spots in Drosophila Although segregation and recombination in mitosis do not occur at the same frequency
Molecular Genetics of. Plant Development STEPHEN H. HOWELL CAMBRIDGE UNIVERSITY PRESS
 Molecular Genetics of Plant Development STEPHEN H. HOWELL CAMBRIDGE UNIVERSITY PRESS Contents Preface A Word on Genetic Nomenclature page xiii xvii 1 Approaches to the Study of Plant Development 1 Pattern
Molecular Genetics of Plant Development STEPHEN H. HOWELL CAMBRIDGE UNIVERSITY PRESS Contents Preface A Word on Genetic Nomenclature page xiii xvii 1 Approaches to the Study of Plant Development 1 Pattern
EPFL signals in the boundary region of the SAM restrict its size and promote leaf initiation.
 Plant Physiology Preview. Published on November 8, 2018, as DOI:10.1104/pp.18.00714 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 EPFL signals in the boundary region
Plant Physiology Preview. Published on November 8, 2018, as DOI:10.1104/pp.18.00714 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 EPFL signals in the boundary region
Arabidopsis Class I KNOTTED-Like Homeobox Proteins Act Downstream in the IDA-HAE/HSL2 Floral Abscission Signaling Pathway C W OA
 This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs,
This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs,
Host-Pathogen Interaction. PN Sharma Department of Plant Pathology CSK HPKV, Palampur
 Host-Pathogen Interaction PN Sharma Department of Plant Pathology CSK HPKV, Palampur-176062 PATHOGEN DEFENCE IN PLANTS A BIOLOGICAL AND MOLECULAR VIEW Two types of plant resistance response to potential
Host-Pathogen Interaction PN Sharma Department of Plant Pathology CSK HPKV, Palampur-176062 PATHOGEN DEFENCE IN PLANTS A BIOLOGICAL AND MOLECULAR VIEW Two types of plant resistance response to potential
A Quantitative and Dynamic Model for Plant Stem Cell Regulation
 A Quantitative and Dynamic Model for Plant Stem Cell Regulation Florian Geier 1,2,7, Jan U. Lohmann 3,4 *, Moritz Gerstung 1,5, Annette T. Maier 3, Jens Timmer 1,6, Christian Fleck 1,7 1 Department of
A Quantitative and Dynamic Model for Plant Stem Cell Regulation Florian Geier 1,2,7, Jan U. Lohmann 3,4 *, Moritz Gerstung 1,5, Annette T. Maier 3, Jens Timmer 1,6, Christian Fleck 1,7 1 Department of
CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON
 PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
PROKARYOTE GENES: E. COLI LAC OPERON CHAPTER 13 CHAPTER 13 PROKARYOTE GENES: E. COLI LAC OPERON Figure 1. Electron micrograph of growing E. coli. Some show the constriction at the location where daughter
Supplementary Materials for
 www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
www.sciencesignaling.org/cgi/content/full/6/301/ra98/dc1 Supplementary Materials for Regulation of Epithelial Morphogenesis by the G Protein Coupled Receptor Mist and Its Ligand Fog Alyssa J. Manning,
Supporting Online Material
 1 Stomatal Patterning and Differentiation by Synergistic Interactions of Receptor Kinases Elena D. Shpak, Jessica Messmer McAbee, Lynn Jo Pillitteri, and Keiko U. Torii Supporting Online Material Material
1 Stomatal Patterning and Differentiation by Synergistic Interactions of Receptor Kinases Elena D. Shpak, Jessica Messmer McAbee, Lynn Jo Pillitteri, and Keiko U. Torii Supporting Online Material Material
Actions of auxin. Hormones: communicating with chemicals History: Discovery of a growth substance (hormone- auxin)
") Hormones: communicating with chemicals History- discovery of plant hormone. Auxin Concepts of hormones Auxin levels are regulated by synthesis/degradation, transport, compartmentation, conjugation. Polar
Hormones: communicating with chemicals History- discovery of plant hormone. Auxin Concepts of hormones Auxin levels are regulated by synthesis/degradation, transport, compartmentation, conjugation. Polar
673 Comparative Genomics of Angiosperm MADS Box Genes Yale University, New Haven, CT. 674 The evolution of plant architecture in Brassicaceae
 673 Comparative Genomics of Angiosperm MADS Box Genes Vivian F. Irish Yale University, New Haven, CT. MADS box genes encode key transcriptional regulators that have been implicated in the control of various
673 Comparative Genomics of Angiosperm MADS Box Genes Vivian F. Irish Yale University, New Haven, CT. MADS box genes encode key transcriptional regulators that have been implicated in the control of various
Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering
 Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering by Valverde et. Al Published in Science 2004 Presented by Boyana Grigorova CBMG 688R Feb. 12, 2007 Circadian Rhythms: The Clock Within
Photoreceptor Regulation of Constans Protein in Photoperiodic Flowering by Valverde et. Al Published in Science 2004 Presented by Boyana Grigorova CBMG 688R Feb. 12, 2007 Circadian Rhythms: The Clock Within
Designer Genes C Test
 Northern Regional: January 19 th, 2019 Designer Genes C Test Name(s): Team Name: School Name: Team Number: Rank: Score: Directions: You will have 50 minutes to complete the test. You may not write on the
Northern Regional: January 19 th, 2019 Designer Genes C Test Name(s): Team Name: School Name: Team Number: Rank: Score: Directions: You will have 50 minutes to complete the test. You may not write on the
Supplementary Materials for
 advances.sciencemag.org/cgi/content/full/2/1/e1500989/dc1 Supplementary Materials for An epidermis-driven mechanism positions and scales stem cell niches in plants Jérémy Gruel, Benoit Landrein, Paul Tarr,
advances.sciencemag.org/cgi/content/full/2/1/e1500989/dc1 Supplementary Materials for An epidermis-driven mechanism positions and scales stem cell niches in plants Jérémy Gruel, Benoit Landrein, Paul Tarr,
Utilizing Illumina high-throughput sequencing technology to gain insights into small RNA biogenesis and function
 Utilizing Illumina high-throughput sequencing technology to gain insights into small RNA biogenesis and function Brian D. Gregory Department of Biology Penn Genome Frontiers Institute University of Pennsylvania
Utilizing Illumina high-throughput sequencing technology to gain insights into small RNA biogenesis and function Brian D. Gregory Department of Biology Penn Genome Frontiers Institute University of Pennsylvania
Major questions of evolutionary genetics. Experimental tools of evolutionary genetics. Theoretical population genetics.
 Evolutionary Genetics (for Encyclopedia of Biodiversity) Sergey Gavrilets Departments of Ecology and Evolutionary Biology and Mathematics, University of Tennessee, Knoxville, TN 37996-6 USA Evolutionary
Evolutionary Genetics (for Encyclopedia of Biodiversity) Sergey Gavrilets Departments of Ecology and Evolutionary Biology and Mathematics, University of Tennessee, Knoxville, TN 37996-6 USA Evolutionary
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 6, 2007 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
23-. Shoot and root development depend on ratio of IAA/CK
 Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Balance of Hormones regulate growth and development Environmental factors regulate hormone levels light- e.g. phototropism gravity- e.g. gravitropism temperature Mode of action of each hormone 1. Signal
Potential Sites of Bioactive Gibberellin Production during Reproductive Growth in Arabidopsis W
 This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs,
This article is a Plant Cell Advance Online Publication. The date of its first appearance online is the official date of publication. The article has been edited and the authors have corrected proofs,
Lecture 4: Radial Patterning and Intercellular Communication.
 Lecture 4: Radial Patterning and Intercellular Communication. Summary: Description of the structure of plasmodesmata, and the demonstration of selective movement of solutes and large molecules between
Lecture 4: Radial Patterning and Intercellular Communication. Summary: Description of the structure of plasmodesmata, and the demonstration of selective movement of solutes and large molecules between
Supplemental Figure 1: Increased Fe deficiency gene expression in roots of nas4x-2
 Supplemental Figure 1: Increased Fe deficiency gene expression in roots of nas4x-2 IRT1, FRO2 and FIT expression levels in roots of the wild-type, nas4x- 1 and nas4x-2, showing that in both nas mutants
Supplemental Figure 1: Increased Fe deficiency gene expression in roots of nas4x-2 IRT1, FRO2 and FIT expression levels in roots of the wild-type, nas4x- 1 and nas4x-2, showing that in both nas mutants
DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Other cellular processes (17%)
 Signal transduction (2%) Development (2%) Other cellular processes (17%)") Fig. 35-24 Other metabolism (18%) DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Unknown (24%) Energy pathways (3%) Cell division and organization (3%) Transport (4%) Transcription
Fig. 35-24 Other metabolism (18%) DNA or RNA metabolism (1%) Signal transduction (2%) Development (2%) Unknown (24%) Energy pathways (3%) Cell division and organization (3%) Transport (4%) Transcription
Plant Structure and Organization - 1
 Plant Structure and Organization - 1 In our first unit of Biology 203 we will focus on the structure and function of the higher plants, in particular the angiosperms, or flowering plants. We will look
Plant Structure and Organization - 1 In our first unit of Biology 203 we will focus on the structure and function of the higher plants, in particular the angiosperms, or flowering plants. We will look
(A) Ethylene (B) Absisic acid (C) Auxin (D) Gibberellin (E) Cytokinin
 Ethylene (B) Absisic acid (C) Auxin (D) Gibberellin (E) Cytokinin") College Biology - Problem Drill 17: Plant Function Question No. 1 of 10 1. Which of the following plant hormones is responsible for phototropism? Question #01 (A) Ethylene (B) Absisic acid (C) Auxin (D)
College Biology - Problem Drill 17: Plant Function Question No. 1 of 10 1. Which of the following plant hormones is responsible for phototropism? Question #01 (A) Ethylene (B) Absisic acid (C) Auxin (D)
CLV3 Is Localized to the Extracellular Space, Where It Activates the Arabidopsis CLAVATA Stem Cell Signaling Pathway
 The Plant Cell, Vol. 14, 969 977, May 2002, www.plantcell.org 2002 American Society of Plant Biologists RESEARCH ARTICLE CLV3 Is Localized to the Extracellular Space, Where It Activates the Arabidopsis
The Plant Cell, Vol. 14, 969 977, May 2002, www.plantcell.org 2002 American Society of Plant Biologists RESEARCH ARTICLE CLV3 Is Localized to the Extracellular Space, Where It Activates the Arabidopsis
Aida, M., Vernoux, X., Furutani, M., Traas, J. and Tasaka, M. (2002). Development 129, On page 3972 of this article, the sentence This
. Development 129, On page 3972 of this article, the sentence This") ERRATUM Roles of PIN-FORMED1 and MONOPTEROS in pattern formation of the apical region of the Arabidopsis embryo Aida, M., Vernoux, X., Furutani, M., Traas, J. and Tasaka, M. (2002). Development 129, 3965-3974.
ERRATUM Roles of PIN-FORMED1 and MONOPTEROS in pattern formation of the apical region of the Arabidopsis embryo Aida, M., Vernoux, X., Furutani, M., Traas, J. and Tasaka, M. (2002). Development 129, 3965-3974.
Axis Specification in Drosophila
 Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
Developmental Biology Biology 4361 Axis Specification in Drosophila November 2, 2006 Axis Specification in Drosophila Fertilization Superficial cleavage Gastrulation Drosophila body plan Oocyte formation
REVOLUTA regulates meristem initiation at lateral positions
 The Plant Journal (2001) 25(2), 223±236 REVOLUTA regulates meristem initiation at lateral positions Denichiro Otsuga 1, Bernadette DeGuzman 2,², Michael J. Prigge 2, Gary N. Drews 1 and Steven E. Clark
The Plant Journal (2001) 25(2), 223±236 REVOLUTA regulates meristem initiation at lateral positions Denichiro Otsuga 1, Bernadette DeGuzman 2,², Michael J. Prigge 2, Gary N. Drews 1 and Steven E. Clark
Characterisation of three shoot apical meristem mutants of Arabidopsis thaliana
 Development 116, 397-403 (1992) Printed in Great Britain The Company of Biologists Limited 1992 397 Characterisation of three shoot apical meristem mutants of Arabidopsis thaliana H. M. OTTOLINE LEYSER*
Development 116, 397-403 (1992) Printed in Great Britain The Company of Biologists Limited 1992 397 Characterisation of three shoot apical meristem mutants of Arabidopsis thaliana H. M. OTTOLINE LEYSER*
Chapter 4 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays.
 Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
Evaluating a potential interaction between deltex and git in Drosophila: genetic interaction, gene overexpression and cell biology assays. The data described in chapter 3 presented evidence that endogenous
a. capture sunlight and absorb CO 2
 BIO 274-01 Exam 1 Name Matching (10 pts) 1. Match each plant part with its function: root c a. capture sunlight and absorb CO 2 for photosynthesis leaves a b. provides support, conducts water and nutrients
BIO 274-01 Exam 1 Name Matching (10 pts) 1. Match each plant part with its function: root c a. capture sunlight and absorb CO 2 for photosynthesis leaves a b. provides support, conducts water and nutrients
BLADE-ON-PETIOLE Dependent Signaling Controls Leaf and Floral Patterning in Arabidopsis
 The Plant Cell, Vol. 17, 1434 1448, May 2005, www.plantcell.org ª 2005 American Society of Plant Biologists BLADE-ON-PETIOLE Dependent Signaling Controls Leaf and Floral Patterning in Arabidopsis Shelley
The Plant Cell, Vol. 17, 1434 1448, May 2005, www.plantcell.org ª 2005 American Society of Plant Biologists BLADE-ON-PETIOLE Dependent Signaling Controls Leaf and Floral Patterning in Arabidopsis Shelley
Bi Lecture 8 Genetic Pathways and Genetic Screens
 Bi190-2013 Lecture 8 Genetic Pathways and Genetic Screens WT A 2X:2A her-1 tra-1 1X:2A her-1 tra-1 Female body Male body Female body Male body her-1(lf) B 2X:2A her-1(lf) tra-1 1X:2A her-1(lf) tra-1 Female
Bi190-2013 Lecture 8 Genetic Pathways and Genetic Screens WT A 2X:2A her-1 tra-1 1X:2A her-1 tra-1 Female body Male body Female body Male body her-1(lf) B 2X:2A her-1(lf) tra-1 1X:2A her-1(lf) tra-1 Female
Analysis of regulatory function of circadian clock. on photoreceptor gene expression
 Thesis of Ph.D. dissertation Analysis of regulatory function of circadian clock on photoreceptor gene expression Tóth Réka Supervisor: Dr. Ferenc Nagy Biological Research Center of the Hungarian Academy
Thesis of Ph.D. dissertation Analysis of regulatory function of circadian clock on photoreceptor gene expression Tóth Réka Supervisor: Dr. Ferenc Nagy Biological Research Center of the Hungarian Academy
SLOW MOTION Is Required for Within-Plant Auxin Homeostasis and Normal Timing of Lateral Organ Initiation at the Shoot Meristem in Arabidopsis C W
 The Plant Cell, Vol. 22: 335 348, February 2010, www.plantcell.org ã 2010 American Society of Plant Biologists SLOW MOTION Is Required for Within-Plant Auxin Homeostasis and Normal Timing of Lateral Organ
The Plant Cell, Vol. 22: 335 348, February 2010, www.plantcell.org ã 2010 American Society of Plant Biologists SLOW MOTION Is Required for Within-Plant Auxin Homeostasis and Normal Timing of Lateral Organ
From basic research to crop improvement. Dirk Inze VIB-UGent Center for Plant Systems Biology
 From basic research to crop improvement Dirk Inze VIB-UGent Center for Plant Systems Biology Oct 2017 The Great Challenge By 2050 70% more food on the same land area Growing world population Climate change
From basic research to crop improvement Dirk Inze VIB-UGent Center for Plant Systems Biology Oct 2017 The Great Challenge By 2050 70% more food on the same land area Growing world population Climate change
Heather Currinn, Benjamin Guscott, Zita Balklava, Alice Rothnie and Thomas Wassmer*
 Online Resources APP controls the formation of PI(3,5)P 2 vesicles through its binding of the PIKfyve complex. Cellular and Molecular Life Sciences Heather Currinn, Benjamin Guscott, Zita Balklava, Alice
Online Resources APP controls the formation of PI(3,5)P 2 vesicles through its binding of the PIKfyve complex. Cellular and Molecular Life Sciences Heather Currinn, Benjamin Guscott, Zita Balklava, Alice
Major Plant Hormones 1.Auxins 2.Cytokinins 3.Gibberelins 4.Ethylene 5.Abscisic acid
 Plant Hormones Lecture 9: Control Systems in Plants What is a Plant Hormone? Compound produced by one part of an organism that is translocated to other parts where it triggers a response in target cells
Plant Hormones Lecture 9: Control Systems in Plants What is a Plant Hormone? Compound produced by one part of an organism that is translocated to other parts where it triggers a response in target cells
Developmental Biology
 Developmental Biology 341 (2010) 95 113 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology Review Twenty years on: The inner workings
Developmental Biology 341 (2010) 95 113 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology Review Twenty years on: The inner workings
Genetic interaction and phenotypic analysis of the Arabidopsis MAP kinase pathway mutations mekk1 and mpk4 suggests signaling pathway complexity
 Genetic interaction and phenotypic analysis of the Arabidopsis MAP kinase pathway mutations mekk1 and mpk4 suggests signaling pathway complexity Shih-Heng Su, Maria Cristina Suarez-Rodriguez, Patrick Krysan
Genetic interaction and phenotypic analysis of the Arabidopsis MAP kinase pathway mutations mekk1 and mpk4 suggests signaling pathway complexity Shih-Heng Su, Maria Cristina Suarez-Rodriguez, Patrick Krysan
Electromagenetic spectrum
 Light Controls of Plant Development 1 Electromagenetic spectrum 2 Light It is vital for photosynthesis and is also necessary to direct plant growth and development. It acts as a signal to initiate and
Light Controls of Plant Development 1 Electromagenetic spectrum 2 Light It is vital for photosynthesis and is also necessary to direct plant growth and development. It acts as a signal to initiate and
Genetic control of branching pattern and floral identity during Petunia inflorescence development
 Development 125, 733-742 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV0153 733 Genetic control of branching pattern and floral identity during Petunia inflorescence development
Development 125, 733-742 (1998) Printed in Great Britain The Company of Biologists Limited 1998 DEV0153 733 Genetic control of branching pattern and floral identity during Petunia inflorescence development
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis
 18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
18.4 Embryonic development involves cell division, cell differentiation, and morphogenesis An organism arises from a fertilized egg cell as the result of three interrelated processes: cell division, cell
Big Idea 1: The process of evolution drives the diversity and unity of life.
 Big Idea 1: The process of evolution drives the diversity and unity of life. understanding 1.A: Change in the genetic makeup of a population over time is evolution. 1.A.1: Natural selection is a major
Big Idea 1: The process of evolution drives the diversity and unity of life. understanding 1.A: Change in the genetic makeup of a population over time is evolution. 1.A.1: Natural selection is a major
Eukaryotic Gene Expression
 Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Eukaryotic Gene Expression Lectures 22-23 Several Features Distinguish Eukaryotic Processes From Mechanisms in Bacteria 123 Eukaryotic Gene Expression Several Features Distinguish Eukaryotic Processes
Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed
 Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed using the Geneious software. Accession numbers of the
Supplementary Figure S1. Amino acid alignment of selected monocot FT-like and TFL-like sequences. Sequences were aligned using ClustalW and analyzed using the Geneious software. Accession numbers of the
Conclusions. The experimental studies presented in this thesis provide the first molecular insights
 C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
C h a p t e r 5 Conclusions 5.1 Summary The experimental studies presented in this thesis provide the first molecular insights into the cellular processes of assembly, and aggregation of neural crest and
Questions for Biology IIB (SS 2006) Wilhelm Gruissem
 Wilhelm Gruissem") Questions for Biology IIB (SS 2006) Plant biology Wilhelm Gruissem The questions for my part of Biology IIB, Plant Biology, are provided for self-study and as material for the exam. Please note that the
Questions for Biology IIB (SS 2006) Plant biology Wilhelm Gruissem The questions for my part of Biology IIB, Plant Biology, are provided for self-study and as material for the exam. Please note that the
AP Curriculum Framework with Learning Objectives
 Big Ideas Big Idea 1: The process of evolution drives the diversity and unity of life. AP Curriculum Framework with Learning Objectives Understanding 1.A: Change in the genetic makeup of a population over
Big Ideas Big Idea 1: The process of evolution drives the diversity and unity of life. AP Curriculum Framework with Learning Objectives Understanding 1.A: Change in the genetic makeup of a population over
Progress in modeling biological development as it bears on SBML
 Progress in modeling biological development as it bears on SBML Eric Mjolsness UC Irvine representing the Computable Plant project www.computableplant.org Three relevant examples Meristem maintenance by
Progress in modeling biological development as it bears on SBML Eric Mjolsness UC Irvine representing the Computable Plant project www.computableplant.org Three relevant examples Meristem maintenance by
Other funding Sources Agency Name: MSU Agricultural Experiment Station /Project GREEEN Amount requested or awarded: 30,000
 FINAL PROJECT REPORT Project Title: Functional genomics of flowering in apple PI: Herb Aldwinckle Co-PI(2): Steve VanNocker Organization: Cornell University Organization: Michigan State University Telephone/email:
FINAL PROJECT REPORT Project Title: Functional genomics of flowering in apple PI: Herb Aldwinckle Co-PI(2): Steve VanNocker Organization: Cornell University Organization: Michigan State University Telephone/email:
Auxin Response Factors Mediate Arabidopsis Organ Asymmetry via Modulation of KANADI Activity W
 The Plant Cell, Vol. 17, 2899 2910, November 2005, www.plantcell.org ª 2005 American Society of Plant Biologists Auxin Response Factors Mediate Arabidopsis Organ Asymmetry via Modulation of KANADI Activity
The Plant Cell, Vol. 17, 2899 2910, November 2005, www.plantcell.org ª 2005 American Society of Plant Biologists Auxin Response Factors Mediate Arabidopsis Organ Asymmetry via Modulation of KANADI Activity
UNIT 6 - STRUCTURES OF FLOWERING PLANTS & THEIR FUNCTIONS
 6.1 Plant Tissues A tissue is a group of cells with common function, structures or both. In plants we can find 2 types of tissues: Meristem Permanent tissues Meristem is found in regions with continuous
6.1 Plant Tissues A tissue is a group of cells with common function, structures or both. In plants we can find 2 types of tissues: Meristem Permanent tissues Meristem is found in regions with continuous
Processes of Evolution
 15 Processes of Evolution Forces of Evolution Concept 15.4 Selection Can Be Stabilizing, Directional, or Disruptive Natural selection can act on quantitative traits in three ways: Stabilizing selection
15 Processes of Evolution Forces of Evolution Concept 15.4 Selection Can Be Stabilizing, Directional, or Disruptive Natural selection can act on quantitative traits in three ways: Stabilizing selection
Cells to Tissues. Peter Takizawa Department of Cell Biology
 Cells to Tissues Peter Takizawa Department of Cell Biology From one cell to ensembles of cells. Multicellular organisms require individual cells to work together in functional groups. This means cells
Cells to Tissues Peter Takizawa Department of Cell Biology From one cell to ensembles of cells. Multicellular organisms require individual cells to work together in functional groups. This means cells
The Science of Plants in Agriculture Pl.Sci 102. Getting to Know Plants
 The Science of Plants in Agriculture Pl.Sci 102 Getting to Know Plants Growth and Development of Plants Growth and Development of Plants Why it s important to have knowledge about plant development. What
The Science of Plants in Agriculture Pl.Sci 102 Getting to Know Plants Growth and Development of Plants Growth and Development of Plants Why it s important to have knowledge about plant development. What
Enduring understanding 1.A: Change in the genetic makeup of a population over time is evolution.
 The AP Biology course is designed to enable you to develop advanced inquiry and reasoning skills, such as designing a plan for collecting data, analyzing data, applying mathematical routines, and connecting
The AP Biology course is designed to enable you to develop advanced inquiry and reasoning skills, such as designing a plan for collecting data, analyzing data, applying mathematical routines, and connecting