Topology of class A G protein-coupled receptors: insights gained from crystal structures of rhodopsins, adrenergic and adenosine receptors
|
|
|
- Annice Davidson
- 5 years ago
- Views:
Transcription
1 Molecular Pharmacology This article has Fast not been Forward. copyedited Published and formatted. on The October final version 22, may 2008 differ as from doi: /mol this version. MINIREVIEW Topology of class A G protein-coupled receptors: insights gained from crystal structures of rhodopsins, adrenergic and adenosine receptors Debarshi Mustafi and Krzysztof Palczewski Department of Pharmacology, Case Western Reserve University, Cleveland, OH, Page 1 Copyright 2008 by the American Society for Pharmacology and Experimental Therapeutics.
2 Running Title: Structures of class A G protein-coupled receptors Correspondence to: Debarshi Mustafi, Department of Pharmacology, School of Medicine, Case Western Reserve University, Euclid Ave., Cleveland, Ohio , USA; Phone: , Fax: , debarshi.mustafi@case.edu Krzysztof Palczewski, Ph.D., Department of Pharmacology, School of Medicine, Case Western Reserve University, Euclid Ave., Cleveland, Ohio , USA; Phone: , Fax: , kxp65@case.edu Document statistics: 26 Number of text pages 1 Table 6 Figures 74 References 119 Words in the Abstract 749 Words in the Introduction 1079 Words in the Discussion Non-standard abbreviations: Metarhodopsin I (Meta I or MI), Metarhodopsin II (Meta II or MII), G protein coupled receptor (GPCR), β 1 -adrenergic receptor (β 1 AR), β 2 -adrenergic receptor (β 2 AR) Page 2
3 ABSTRACT: Biological membranes are densely packed with membrane proteins that occupy about half of their volume. In almost all cases, membrane proteins in the native state lack the higher-order symmetry required for their direct study by diffraction methods. Despite many technical difficulties, numerous crystal structures of detergent solubilized membrane proteins have been determined that illustrate their internal organization. Among such proteins, class A G protein-coupled receptors (GPCRs) have become amenable to crystallization and high resolution x-day diffraction analyses. The derived structures of native and engineered receptors not only provide insights into their molecular arrangements, but also furnish a framework for designing and testing potential models of transformation from inactive to active receptor signaling states and for initiating rational drug design. Page 3
4 Introduction G protein-coupled receptors (GPCRs) comprised of seven transmembrane helices are the largest superfamily of proteins in the human body. Disruptions of most individual GPCR genes in mouse models is not lethal or developmentally harmful because genes encoding receptors responding to the same ligand(s) are highly redundant, and/or encode regulatory proteins for physiological processes that can be controlled by different cellular mechanisms in case of failure. GPCRs, however, modulate almost all physiological processes because they are ubiquitous and largely located in plasma membranes where they are readily accessible to endogenous signaling molecules making them extremely attractive drug targets. The human genome sequence (Lander et al., 2001; Venter et al., 2001) indicates that of the ~35,000 human genes, ~720 encode GPCRs, of which ~ 400 are expected to be potential drug targets (Kroeze et al., 2003). GPCR agonist and antagonist drugs have been successfully used to treat patients with a broad spectrum of diseases owing to the large diversity of involved receptors (Drews, 2000; Wise et al., 2002). For example, common human disorders associated with vision (involving rhodopsin mutations)(menon et al., 2001), the cardiovascular system (caused by β 1 -adrenergic receptors)(drake et al., 2006), asthma (attributable to β 2 -adrenergic receptors)(kawakami et al., 2004), and strokes and cerebral hypoperfusion (altered adenosine A 2A receptor function in accompanying heart disease) all involve aberrant GPCR signaling. Identifying the pathophysiological roles of these potential drug targets is fundamentally dependent on understanding their inherent structures and threedimensional ligand-protein interactions (Klabunde and Hessler, 2002). Detailed structural interactions between ligands and GPCRs are largely unresolved because of the inherent difficulty of membrane protein crystallization (Loll, 2003) despite new high-throughput technologies designed to limit the amounts of sample required (Hansen et al., 2006; Hansen et al., 2002; Li et al., 2006). Elucidation of the first mammalian GPCR structure of native bovine rhodopsin (Palczewski et al., 2000) and subsequent higher resolution structures (Li et al., 2004; Okada et al., 2004; Stenkamp, 2008) have opened the way to probe ligand binding sites and assess structure-function relationships for these membrane proteins (Hubbell et al., 2003; Lu et al., 2002; Park et al., 2004). The newest x-ray defined coordinates encompass all amino acids and posttranslational modifications (Table 1). A mutant recombinant bovine rhodopsin structure solved at lower resolution (3.4 Ǻ) than native rhodopsin (2.2 Å) also has been reported Page 4
5 (Standfuss et al., 2007)(Table 1), and it highlights the possibility of using recombinant proteins for structural studies, as developed recently for adrenergic and adenosine receptors. Moreover, the structure of isorhodopsin was solved in which the native 11-cis-retinal of rhodopsin is replaced with the analog, 9-cis-retinal (Nakamichi et al., 2007; Nakamichi and Okada, 2007). No significant structural differences were noted between rhodopsin and isorhodopsin. The x-ray defined structure of bovine rhodopsin initiated homology modeling of GPCRs (Filipek et al., 2003b; Patny et al., 2006) to understand the structures of other functionally important GPCRs and relate them for rational drug design (Becker et al., 2003). Despite these efforts, the above methods failed to predict models and possible ligands efficiently for therapeutic purposes (Deupi et al., 2007). The only structural features common to all GPCRs are their seven transmembranespanning α-helical segments connected by alternating intracellular (C-) and extracellular (E-) loops, with the amino terminus located on the extracellular side and the carboxyl terminus on the intracellular side. Significant sequence homology is found, however, within several subfamilies of GPCRs engaged in redundant or unique functions (Gether, 2000). These typically are classified by the presence of specific motifs, ligand sizes, relationships to a reference receptor, and other criteria. The three major GPCR subfamilies in vertebrates include those related to rhodopsin, adrenergic and adenosine receptors, totaling ~280 GPCRs (Kroeze et al., 2003). These receptors can be classified as those that show a highly conserved sequence homology of an Asp-Arg-Tyr motif on the intracellular side of the C-III loop (class A); the secretin-receptor family (class B); and those related to metabotropic glutamate receptors (class C)(Gether, 2000). Recently, x-ray crystallography of structural forms of native rhodopsins, mutant β 1 -adrenergic, and engineered β 2 -adrenergic and A 2A adenosine receptors have been solved at high resolution. This review focuses on the elucidation of six new class A GPCR structures and how their structural features differ slightly from those existing in native bovine rhodopsin. Different motifs specific to each new structure are featured that confer its unique ligand specificity and activity. Also addressed are structural artifacts attributable to the use of mutations and fusion proteins for x-ray crystallography and the future implications of structural work for cellular signaling mechanisms and drug design. Different photointermediates of rhodopsin Page 5
6 Rhodopsin is a GPCR expressed in rod retinal photoreceptors that plays a key role in vision by converting light into a cascade of biochemical transformations called phototransduction (Filipek et al., 2003a; Palczewski, 2006). Rhodopsin was the first crystallized GPCR (Okada et al., 2000; Palczewski et al., 2000), and subsequent work resulted in crystallization of rhodopsin photointermediates that improved understanding of the chromophore transformation cycle. The membrane embedded chromophore contained in rhodopsin is 11-cis-retinal. This is covalently bound to the inactive opsin protein by a Lys residue (Lys 296 ) in helix VII via a protonated Schiff base. Upon absorption of a photon, isomerization of this chromophore to an all-trans conformation induces changes in the opsin structure, ultimately converting it from an inactive to an activated signaling state. The last form of this receptor, known as metarhodopsin II (MII, meta II, or R * ), relays the activating changes to the retinal G protein (guanine-nucleotide-binding protein), transducin (G t ), that in turn initiates the series of biochemical reactions culminating in a neuronal signal. As the chromophore converts to the all-trans conformation during photoactivation, rhodopsin progresses through a series of photointermediates. The first structurally characterized photointermediate, bathorhodopsin, thermally relaxes to the blue shifted intermediate bathorhodopsin (BSI), followed by lumirhodopsin and then metarhodopsin I (MI or meta I). In MI, the all-trans-retinal chromophore remains bound to opsin. However, during the MI to MII transition, the all-trans-retinylidene Schiff base becomes deprotonated. MII, the signaling state capable of G protein activation, ultimately decays to form free all-trans-retinal and opsin. Interestingly, it has been shown that there are two forms of MI and MII, MIa/MIb and MIIa/MIIb, respectively, both in a ph-dependent equilibrium; MIIb is the only MII partner capable of activating G t (Okada et al., 2001; Ridge and Palczewski, 2007; Schertler, 2005). Because photoactivation of rhodopsin involves formation of a series of reaction intermediates having different shapes and dissimilar retinal ligands, different forms of rhodopsin crystals should be ideal for studying the mechanism of GPCR activation. With a high light absorption coefficient, the intrinsic chromophore of rhodopsin is a sensitive indicator of conformational changes because of its sensitivity to minor changes in the first, second and even the third environmental shell surrounding the chromophore binding pocket. This property is harnessed by nature that uses binding of an identical chromophore to different opsins to regulate the λ maximum absorption, or so-called opsin shift, to provide a molecular mechanism for color Page 6
7 vision. Because protonated retinylidene interacts with its counterion, minimal changes in the distance between this pair can produce large changes in the λ maximum, irrespective of changes in the overall protein structure (Figure 1). This hypothesis is well supported by rhodopsin mutants with engineered disulfide bridges connecting helices to prevent large conformational changes that both spectrally and in G protein assays achieve a MII state (Yu et al., 1995). Thus, identification of rhodopsin intermediates by their spectral properties alone constitutes a fundamental problem if changes in the λ maxima of these photointermediates are considered indicative of alterations in protein conformation rather than just a reflection of chromophore status. Three-dimensional structures of bathorhodopsin and lumirhodopsin were obtained after trapping these photolyzed states at low temperatures while that of MI was resolved by using electron crystallography of two dimensional crystals (Nakamichi and Okada, 2006a; Nakamichi and Okada, 2006b; Ruprecht et al., 2004). Recently, a structure of the deprotonated photoactivated state of rhodopsin, resembling MII, has also been elucidated (Salom et al., 2006b). The first intermediate trapped following light absorption by rhodopsin is bathorhodopsin; this occurs within a few hundred femtoseconds at room temperature. X-ray diffraction was used to calculate the difference in densities corresponding to the retinal. The overall residual structural changes are slight, but retinal does undergo a small displacement in the β-ionone ring defined by the binding pocket that has room on the extracellular side of the ring (Figure 1). Studies suggest that the 11-cis-retinal binding pocket is designed to achieve a dual function, i.e. retain retinal as an inverse agonist in the dark and allow its efficient photoconversion to an agonist upon exposure to light (Nakamichi and Okada, 2006a). Bathorhodopsin crystals were illuminated with visible light and warmed, resulting in a gradual conversion to lumirhodopsin, the second trappable intermediate. Retinal in lumirhodopsin assumes an all-trans structural conformation. Compared with retinal in bathorhodopsin, there is relaxation of the curved polyene chain. This process is proposed to be the key step in transferring stored photon energy to the surrounding protein moiety by converting the 11-cis-retinylidene inverse-agonist to an all-trans agonist (Nakamichi and Okada, 2006b) (Figure 1). An equilibrium is formed between the later photointermediates, MI and MII. MII corresponds to the fully activated receptor, which binds to and activates the heterotrimeric G t. MI Page 7
8 was studied by freeze-trapping and electron microscopy. Density maps revealed that MI formation does not involve large movements or rotations of rhodopsin s helices, but rather, as noted with the other photointermediates, conformational changes localized around the chromophore binding pocket. At this stage, photon energy is fully transferred to the proteinchromophore complex and the excess of energy dissipated as oscillation vibrations (Ruprecht et al., 2004). The subsequent transformation from MI to MII is driven by enthalpy. The inherent instability of rhodopsin in its photoactivated deprotonated state required development of a purification protocol and crystallization conditions that was capable of withstanding photoactivation (Salom et al., 2006a). This advance permitted the growth of ground state crystals, albeit at lower resolution (4.1 Ǻ) than other rhodopsin structures, that upon exposure to light transformed rhodopsin into a photoactivated deprotonated intermediate resembling the MII biological state. Unlike the other rhodopsin structures, the photoactivated structure did not have residues V230-Q238, K311-F313, D330-A348 resolved (Table 1) as a result of low resolution and/or flexibility of structural regions. Nonetheless, fine changes were noted in this photoactivated structure (Figure 2A, 2B). The x-ray crystallographic data elucidated that photoactivation primarily causes relaxation of the C-III loops, before the chromophore is hydrolyzed from the binding site and rhodopsin is regenerated from opsin by freshly synthesized 11-cis-retinal (Travis et al., 2007). A higher resolution structure of this intermediate is needed. Crystallographic snapshots of these photointermediates help elucidate the process of chromophore turnover and indicate that the entire activation process entails subtle changes in the overall structure of rhodopsin. The importance of minimal changes found in the photoactivated structures is further highlighted by the potential for dimers of rhodopsin to explain its physiological function. Non-physiological dimers of rhodopsin can be found in its first 3D crystals, where the two subunits are related by a rotation of 180 about an axis in the plane of the membrane that appear to be induced by experimental crystallization conditions (Okada et al., 2001; Palczewski et al., 2000; Teller et al., 2001). However, the new crystal form of the photoactivated structure unveiled a possible physiologically relevant dimer rhodopsin interface. The photoactivated crystal reveals that the dimer is stabilized by a series of intermolecular contacts previously observed in other 3D crystals but rotated by 180 o around a hydrophobic center (Salom et al., 2006b). These results are consistent with data obtained by atomic force Page 8
9 microscopy and molecular modeling (Filipek et al., 2004; Fotiadis et al., 2003; Liang et al., 2003; Liang et al., 2004). Moreover, the finding of dimeric rhodopsin crystals offers a potential explanation for its higher-order organization in native membranes and provides a template to study GPCR activation by crystallography (Muller et al., 2008). Structural similarity between the activated and ground states shows that rhodopsin is a good template for homology models of other GPCRs. We think that an urgent priority now includes the elucidation of the structural complexes of photoactivated rhodopsin with its partner proteins, G t, rhodopsin kinase (GRK1) and arrestin. These structures will provide a truer physiologic perspective of how GPCRs achieve their signaling conformation(s). The complexity of these proteins, especially the G proteins, as well as the instability and heterogeneity of such complexes will make these investigations extremely challenging. Unfortunately, domain approaches are not suitable for GPCR studies because their entire structures form one signaling domain. As illustrated by work with peptides encompassing different regions of GPCRs and interacting proteins, fragments of these receptors partner proteins inadequately mimic the native proteins interactions and their thermodynamic properties. Comparison of invertebrate and vertebrate structures: squid vs bovine rhodopsin In contrast to vertebrate vision, wherein signal transduction is mediated by the second messenger cyclic GMP, invertebrate phototransduction employs an inositol-1,4,5-triphosphate signaling cascade in which a G q protein is stimulated by photoactivated rhodopsin (Terakita et al., 1998). Understanding the invertebrate visual transduction process was enhanced by discovery of the x- ray defined structure of squid rhodopsin (Murakami and Kouyama, 2008). The large C-terminus of squid rhodopsin was proteolitically removed and the M1-N8 residues were not resolved (Table 1). Invertebrate rhodopsin forms a stable pigment with either 11-cis- or all-trans retinylidene, so resolution of these structures constitutes an initial step in learning about the critical determinants protecting the chromophore from hydrolysis. Comparison of squid and bovine rhodopsin in a tetragonal P4 1 crystal (Okada et al., 2004) revealed that the most notable difference is in the C-III loop, a difference attributable to the extra sequence of squid rhodopsin in this region (Figure 2C, 2D). The C-III loop of bovine rhodopsin takes on different conformations in a different crystal forms (Li et al., 2004; Stenkamp, 2008). Extensions of Page 9
10 helices V and VI into the cytoplasmic medium are likely important structural motifs for specifying the coupling mode with G q. Thus, squid rhodopsin provides insights into the differences in rhodopsin signaling mechanisms between vertebrates and invertebrates. Understanding chromophore exchange in vision: opsin vs. rhodopsin Native bovine opsin, an inactive form of rhodopsin lacking the chromophore, was crystallized (Park et al., 2008a; Scheerer et al., 2008) by optimizing the selective extraction of rhodopsin from rod cell disc membranes (Okada et al., 2000; Okada et al., 1998). This methodology enabled crystallization without any modification of the protein that might cause structural distortions. Structural examination of this opsin reveals only slight changes relative to rhodopsin for transmembrane helices I to IV. The most obvious differences are found in the region of transmembrane helices V to VII; these are especially prominent at the cytoplasmic ends of these helices and cause rearrangement of the C-II and C-III loops (Figure 3A, 3B). Furthermore, the ionic lock is broken in the opsin structure such that the rearranged cytoplasmic ends of transmembrane helices five and six are locked by two new interactions. Lack of the interacting prosthetic group of 11-cis-retinal causes a few distinct structural alterations in the retinal-binding pocket. Part of the space occupied by the β-ionone ring is filled in opsin by the side chain of Trp 265. Also, lack of stabilizing residues to hold the polyene chain of the retinal in place causes helix III and the C-III loop to move slightly away from transmembrane helices V to VII. This allows the retinal-binding pocket to become wider towards the retinal attachment site of Lys 296 (Park et al., 2008a; Scheerer et al., 2008). The C-terminus of the opsin structure is not resolved from residues P327-A348 (Table 1). However, a complex between opsin and the C-terminal peptide derived from α-subunit of G protein transducin optimized for tight binding was recently reported (Scheerer et al., 2008) (Table 1). In our opinion, further experiments are essential to show regeneration of opsin crystals back to rhodopsin with 11-cis-retinal chromophore because opsin is inherently unstable in detergents and unable to rebind chromophore (Buczylko et al., 1996; McKibbin et al., 2007). This approach would also provide conclusive evidence that the crystal form reported is of true biologic relevance. The previously studied structure could represent opsin or a more stable conformation of opsin that has passed the stage of regenerability. Page 10
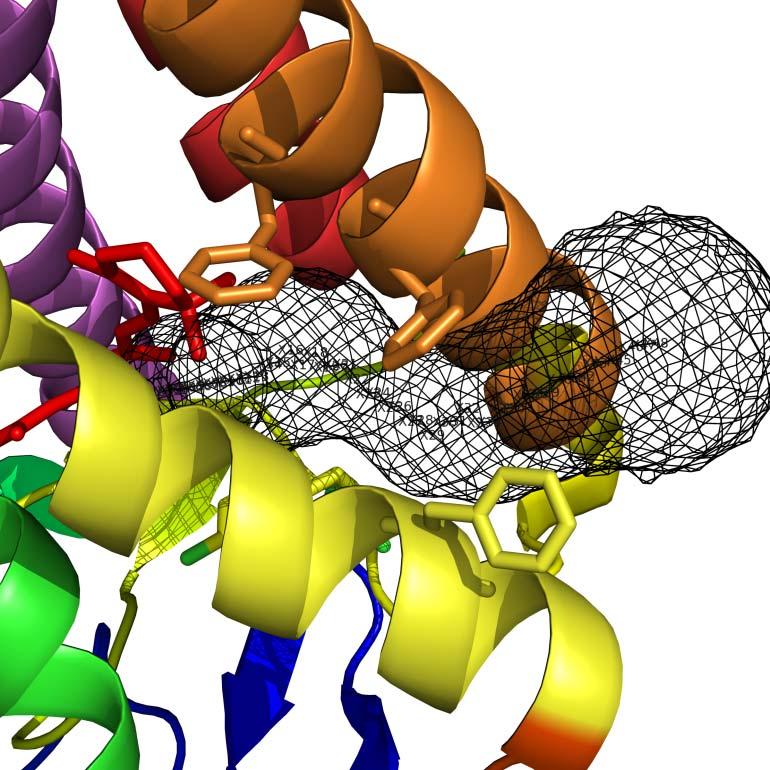
11 The two different openings of the retinal-binding site in opsin suggesting different retinal entrance and exit routes support the hypothesis that all-trans-retinal exits via a pathway differing from the entrance route and remains bound to an exit site (Schadel et al., 2003) with the opening between helix I and helix VII serving as a possible release structure for all-trans-retinal. The proposed uptake route between the extracellular ends of helix V and helix VI may be a general route of ligand uptake among GPCRs as evidenced by our current simulations and past chemical work. Amine ligands for the β 2 -adrenergic receptor exhibit structural similarities to retinal and the aromatic carbazole ring of the inverse agonist, carazolol, could enter the ligand binding pocket at the opposite side of the E-III loop, between helix V and helix VI (Park et al., 2008a; Scheerer et al., 2008). Studies before the opsin structure was elucidated (Schadel et al., 2003) suggested the existence of a tunnel for retinal to traverse, but it was not until the opsin structure was solved that tunneling of the ligand could be confidently deduced. Use of the online software Caver (Petrek et al., 2006) allowed elucidation of the tunneling pathway for retinal into the binding pocket to covalently bind to Lys 296 (Figure 4). This shows two distinct sites, validating the hypothesis that the ligand enters and exits through different parts of the protein. The larger opening between helix V and helix VI (Figure 4) is logical for the uptake of 11-cis-retinal while an exit route for all-trans-retinal through the opening between helix I and helix VII also is realistic owing to its more sterically constricted tunneling configuration. The β-adrenergic and adenosine receptors β 1 -adrenergic receptor vs rhodopsin Rhodopsin and adrenergic receptors belong to class A, the largest and most studied of all GPCR classes. In a crystallized mutant form of the β 1 -adrenergic receptor in complex with the highaffinity antagonist cyanopindolol, six residues were mutated (R68S, M90V, C116L, Y227A, A282L, F327A, F338M, and C358A), three regions were deleted (D2-S32, C244-R271, and A368-A496), and large portions of the structural M1-G2, A33-Q39, R239-R243, A272-M283, and P360-L367 regions were not resolved (Table 1). Despite the overall structural plasticity, the resulting modified structure has some parts that differ from those of native rhodopsins. In all GPCRs, C-II and C-III loops are believed to have a role in the binding, selectivity, and activation of G proteins with the C-II loop being important for the strength of the interaction and the C-III loop for specificity (Wong and Ross, 1994). The difference in the conformation of C-II loop also Page 11
12 is important because this region is highly conserved between β 1 - and β 2 -adrenergic receptors, although it is poorly conserved between these structures and rhodopsin. In a mutated form of the turkey β 1 -adrenergic receptor (Figure 5), the C-II loop forms a short α-helix parallel to the membrane surface whereas in both the mutant β 2 -adrenergic receptor structures and in rhodopsin this loop is in an extended conformation (Warne et al., 2008). The C-III loop is absent in the β 2 - adrenergic receptor-t4 chimera crystal structure and sequestered by the Fab in the other β 2 - adrenergic receptor structure. Destruction of important structural motifs like this probably has compromised the utility of these structures because they do not represent native conformations. Crystallographic artifacts also may change structural insights into ligand binding, and thus adversely affect rational drug design. This alteration is reflected by the mutated turkey β 1 - adrenergic receptor structure to which the natural agonists noradrenaline and isoprenaline bound more weakly by factors of 2,470 and 650, respectively, than to the WT form (Warne et al., 2008). β 2 -adrenergic receptor vs rhodopsin The overall structure of the β 2 -adrenergic receptor with its partial inverse agonist, carazolol, is similar to that of rhodopsin and the β 1 -adrenergic receptor, with seven transmembrane helices and an eighth helix that runs parallel to the cytoplasmic face of the membrane (Figure 6) (Cherezov et al., 2007; Hanson et al., 2008). However, the β 2 -adrenergic receptor was highly engineered with E122W, N187E, Q231-S262 (replaced by amino acids of T4 lysozyme) mutated in the protein, C1054T and C1097A mutated in the T4 lysozyme, and C-terminally Y366-L413 truncated. Moreover, M1-R28 and R343-G365 were not resolved in this structure (Table 1). Thus, we think that any detailed comparison with rhodopsin should be considered with caution. Even so, the structurally conserved helices provide a common core present throughout class A GPCRs, whereas the variable helices confer binding-site plasticity with a resulting architecture capable of binding a large spectrum of ligands. Specifically, and in contrast to rhodopsin, the β 2 -adrenergic receptor has a more open structure. Both forms of the fusion receptors show that the largest difference is in helix I, which is relatively straight and lacks the Pro-induced kink found in rhodopsin; this feature may meet the need for an accessible ligand binding site in the β 2 -adrenergic receptor. Differences in the arrangement of the C- loops of the transmembrane segments of the β 2 -adrenergic receptor and rhodopsin may provide structural Page 12
13 insights into basal receptor activity, given that rhodopsin has no detectable basal activity in contrast to the relatively high basal activity of the β 2 -adrenergic receptor (Rasmussen et al., 2007). However, the β 2 -adrenergic receptor-t4 chimera crystal displayed cholesterol mediated dimeric contacts (Hanson et al., 2008) in a similar orientation as discussed for rhodopsin. Information from the Fab/mutant β 2 -adrenergic receptor complex also provides a global understanding of receptor topology, but the structure is less informative with regard to the precise receptor topology related to physiologic ligand interactions since its crystallization involved similar truncations (Y366-L413) and unresolved residues M1-M36, K60, A91-E107, S165-A202, Q243-K263, V292-L310, A349-G365 (Table 1) as well as adaptation of the C-III loop to bind the Fab antibody fragment. Adenosine receptor vs rhodopsin Like the β 2 -adrenergic receptor, the human A 2A adenosine receptor was crystallized after applying a T4L fusion strategy wherein most of the third cytoplasmic loop is replaced with lysozyme and the carboxyl-terminal tail (A317-S412) is truncated; the resulting structure also revealed that M1-P2, Q148-S156, Q311-A316 were unresolved (Table 1). Despite these similarities, the adenosine receptor has a phospholipid bound in the cholesterol consensus motif. There is also evidence that may suggest that the fusion protein may have shifted the structure toward the activated state, as evidenced by an increased affinity for the subtype-selective agonist CGS21680 as compared to the wild type construct. The helical arrangement is similar among the human β 2 AR, turkey β 1 AR, and squid/bovine rhodopsin structures determined to date. However, the binding pocket of the A 2A adenosine receptor is shifted closer to helices VI and VII, compared to the position of the retinal and carazolol binding pocket in the vicinity of helices III, V, and VI. Also, the E-II loop of the adenosine receptor differs considerably from its counterparts in β 2 AR, β 1 AR, and rhodopsin, in that it lacks any prominent secondary structural element and possesses three disulfide linkages with the E-I loop. This contributes to an extensive disulfide bond network that forms a rigid, open structure, exposing the ligand binding cavity to solvent. The ZM ligand in the crystallized adenosine receptor occupies a significantly different position than retinal and amine ligands, lying perpendicular to the membrane plane. This new structure indicates that the binding pocket in GPCRs can vary in position and Page 13
14 orientation, permitting greater receptor diversity and ligand selectivity. We believe that this finding further highlights the need for high-resolution GPCR structural determinations because the results of molecular modeling deviate significantly from the empiric data now available for the adenosine receptor. GPCR structure and pharmacology Although classical drug screening programs have been successful, more structural knowledge is needed for optimal drug design and improved therapy. The paucity of structural knowledge about GPCRs has severely limited the application of structure-based drug design (Lundstrom, 2006). Even though >30% of all marketed therapeutics act on GPCRs (Hopkins and Groom, 2002), these drugs target only ~30 members of this class so there is enormous potential to exploit the remaining family members, including the >100 orphan receptors (Civelli, 2005) for which no existing ligands have yet been identified. Even when a GPCR has high sequence homology and algorithms are used to predict its overall structure and possibly the location of the small-molecule binding site within the transmembrane domain with some accuracy, small differences in the exact helical arrangement may greatly affect the shape of the small-molecule binding site. For the many class A GPCRs that have a low sequence homology with bovine rhodopsin, the only native ligand bound GPCR structure known to date, homology models cannot accurately predict helical topography, and even less so, the topology of the small-molecule binding site (Schlyer and Horuk, 2006). This problem further highlights the need for native ligand bound states of GPCRs to improve drug design. The structures of β 1 - and β 2 -adrenergic receptors, although informative on a global scale, lack this native binding property, and the new fusion A 2A adenosine receptor illustrates how empiric data is in stark contrast to the predictive molecular modeling data concerning the actual ligand binding pocket. Improved expression and purification methods are badly needed to obtain high resolution structures in their native conformations for optimal drug design. What have we learned about GPCRs from their structures? Several additional points are worth mentioning. (1) It is remarkable how similar the overall structures of rhodopsin, adrenergic and adenosine receptors really are. This fact is easily Page 14
15 forgotten when relatively minor differences are featured in each new elucidated structure. For all rhodopsin structures, root mean square displacements (RMSDs) for transmembrane regions are within 1.8 Å of each other, and even when the adrenergic receptors are compared to rhodopsin, these displacements are Å for β 2 -adrenergic receptor and for β 1 -adrenergic receptor - closer to rhodopsin than to each other (D. Lodowski, unpublished). This close superimposeability occurs, in large part, because of the similar structures of interacting proteins such as the G proteins, receptor kinases (GRKs) and arrestins. Despite differences in overall sequence homology among GPCRs, this natural phenomenon allows the vast majority of these receptors to present a similar topography. Preservation of only a few essential features is likely to be required for the activation process and engagement with G proteins (Mirzadegan et al., 2003). (2) A large number of compounds can bind to each of the GPCRs, yet only a portion of this ligand binding results in receptor activation and coupling with partner proteins. Moreover, among these agonist-stimulated activations, full or partial activity can be achieved. These facts indicate that changes on the cytoplasmic surface lead to differing efficacy of these ligands in activating a particular GPCR. Overall small changes within 2-6 Å observed for rhodopsins and anticipated for other receptors suggest that such subtle changes either directly lead to different receptor conformations, or indirectly affect the extent of protonation/ deprotionation of key residues (like these in the DRY motif region) on the cytoplasmic surface that are responsible for the efficacy of G protein coupling. For photoactivated rhodopsin, meta-stable photointermediates of the activated receptor can be differentiated in part from one another based on their protonation state. The spectrally and functionally distinct MII intermediate capable of activating the heterotrimeric G protein differs from its precursor photointermediates by taking up a proton from the bulk solvent that confers increased conformational flexibility (Salom et al., 2006b). (3) The inherent flexibility of GPCRs permits conformational changes to be triggered by ligand binding. Energy for such a transformation due to ligand binding is typically between 8-12 kcal/mol. Obviously, only a small fraction of this energy is required for activation, and thus large rigid body movements are very unlikely to occur as they are not supported by the energetics of this process. These examples show that even though embedding proteins in a lipid bilayer imposes several restrictions on their conformation and movement, membrane proteins still retain considerable flexibility and mobility that are intimately connected to their function. For class A GPCRs, movement of the kind imposed by the Pro residue in helix VI could be the only Page 15
16 significant movement for part of this helix during activation. Stevens noted that the organization of helices in inactive forms of GPCRs differ slightly from each other (Jaakola et al., 2008). It is temping to speculate that these slight differences in helix organization are not sufficient for activation of these receptors. Thus, how does the allosteric activation of GPCRs take place? (4) Water molecules could be involved in the activation process by an already mentioned protonation/deprotonation mechanism.. Many candidate water molecules have already been identified in GPCRs so discussion of this critical point in GPCR signaling will be the focus of additional theoretical and experimental work. With the lack of major movements along GPCR s helices, water appears to be a logical choice to transmit the signal from the ligand binding site to the cytoplasmic surface through realigning the position of amino acid rotomers along pathways connecting these two sites. (5) Finally, membrane proteins often oligomerize because exposure of hydrophobic regions to water incurs a large energetic penalty. Moreover, immobilization and conformational constraints imposed on these proteins by their oligomerization would provide a stabilizing influence. Both oligomerization and dissociation to monomers can be described by two biophysical concepts that appear largely adequate to explain both the size and formation dynamics of these structures, as has been shown for syntaxin 1 oligomers (Periole et al., 2007). A detailed analysis of oligomerization and its energetics has recently been presented (Muller et al., 2008). Even transient self-association of membrane proteins will guarantee that sufficient time for catalysis or signaling is achieved. Studies of GPCR oligomerization have generated much passionate discussion (Chabre et al., 2003; James and Davis, 2007; Park et al., 2004). However, an overwhelming amount of data accumulated over the years support the concept that GPCRs function as oligomeric rather than monomeric receptors (Milligan, 2004; Park et al., 2004; Terrillon and Bouvier, 2004). This concept is slowly gaining recognition, even in structural studies of GPCRs that provide the intellectual basis for pharmacologically relevant allosteric regulators (Park et al., 2008b). Conclusions Progress in elucidating the structures of membrane-bound GPCRs has been astonishing despite considerable technical difficulties. X-ray diffraction of GPCR crystals provides a static snapshot of their structure that allows simultaneous imaging of the involved structural amino acids. It can be argued that GPCR crystals do not mimic native structures as they form in high concentrations Page 16
17 of the precipitant resulting in internal crystal contacts that could deform the structure and prevent conformational alterations in response to changes in ligand occupancy. This concern is likely valid to some degree, even though the precipitants were selected because they are non-denaturing and crystals would never form from an assembly of differing protein conformations. Moreover, the examined structures are well hydrated (~70%) and the protein concentration within the crystals of rhodopsin is between 9-14 mm, not much above the 5 mm found in the native state (Nickell et al., 2007). Furthermore, detergent and residual or added lipids do provide a milieu sufficient in many cases to maintain the functional properties of GPCRs. Undoubtedly though, there is a critical need to correlate x-ray crystallographic findings with the cellular properties of these receptors. Even though crystallography of membrane proteins is labor intensive and time consuming, there is no obvious substitute yet. For a receptor with dimensions of 75 Å x 45 Å x 30 Å and conformational changes within 1-6 Å, other methods with errors within these ranges or possible artifacts exceeding 10 Å derived from chemical modifications of examined proteins may not be too informative. There also is the concern that structural information gained from extensively engineered GPCRs may or may not be useful for rational drug design where the native human protein structure is critically needed. ACKNOWLEDGEMENTS We would like to thank Dr. Leslie T. Webster Jr. (Case Western Reserve University) for valuable comments on the manuscript. Page 17
18 References KD ^ ^ D z E ^ dddd D d 'WZ K e d ddd ded : ^ : Z< W < deee D : ded dd ddedd ddedd D Z ^, dddd E dde eeed dd dd s Z D, D Z ^'& d &^ < d^,: < W t t/ < < ^ Z ddde, d ' ^ dde dedd ddde dded K dddd 'WZ d W ^ de d dd de y E > Z D> D : W > ddde ^ ' / d D e dd eed eee Dd ^ ^< > Z: ddde d ' Z ee e ded ded : dddd ^ dee dded deed deed & ^ < < & > z ^ W < dddd ' ' W W ^ d e ede ede & ^ ^ Z d W < dddd ' W Z W ed edd eee & ^ d W < ^ Z dddd d ' Z ^ dd ded dee & > z & ^ ^ W < dddd Z E ddd eede dde dde ' h dddd h ' Z dd d ed ddd, > ^ :D Y ^Z ddde : ^ dde dd dddd dddd, > ^ :D Y ^Z dddd W E ^ h^ ee de deddd dedde, D s ' Dd Z : sw z s : < W ^ Z ddde d e d ^ de e eee edd, > ' Z dddd d E Z d e ede edd, t>, D <,' dddd Z D W ddd ded W / ^ Page 18
19 : sw ' Dd, D s z > :Z / W ^ Z ddde d d e ^, d Z ^ : :Z ^: ddde Z Zd 'WZ E d e edd < E D <, ^ &, dddd d, d d :W ^ ed d dde dde < d, ' dddd ' d dd ede edd < t< ^ : Z > dddd ' : dde W dd deee deee > ^ > >D dddd / E dde eedd eed edd > : W D s ^ '&y dddd ^ :D ddd d ddde ddde > > D & Y d s >> d : / Z& ddde E W E ^ h^ddd dd deddd dedde > z & & ^ ^ W < dddd K ' : dee dd ddedd ddeed > z & D d D D & ^ ^ W < dddd Z : dee de dedee dedee > W: dddd D :^ ddd d ddd ddd > > ^ :t, dddd ^ d W ^ dd d ddd dde > < ddde > ' W W ^ e d ded ded D < & E : Z W: <,' t W s W: ddde K : ded d ddde dddd D ^d, D ^ dw dddd Z ^ W Z ed d dede deee D ' dddd ' D ee d d e D d ' & ^ W < dddd ^ ' dd dd dede deee D : t E W < ddde s W Z ed d dd ee D D < d ddde E ddd eded ded hddd E, s K d ddde W e ed dd >dde dde E, K d ddde / dd de dded dded Page 19
20 E, K d ddde > W E ^ h^ddd dd ddede ddedd E, K d ddde y e W ed d ddd ddd E ^ W W^ t W < ddde d d : dee d ede edd K d KW W <, <W dddd d ^ de d dde ddd K d > d / & ^ Z W < dddd y Z : ddd d ed ed K d ^ D E D W s dddd d d d :D ddd d ded ded K d d < < d deee, W W ee d ded dee W < ddde ' ed edd eee W < < d, d D, & > d / d K d ^ Z z D D D dddd ' ^ dee dded ede edd W :, ^ W, <W,t KW ddde ' E eddd ded dee W W^, & ^ t :t W < dddd K ' W dd dd ddedd ddede W W^, > d W < ddde ' Z W d de dde ddd W Ws D ddde, ' D dd dd deee deed W y, d D ^: ^ dw ddde 'W Z ^ ^ D : ^ dde dd dddde ddddd W D K D W < W < : : ddde sz e Z ^'&,: Z D < d^ d &^ W D Z szw ^ Z & Z& ^ '&y t t/ < < ddde d ' E ddd edee ded hded Z < W < ddde s ^ ' : ded dd edee eddd Z :: D d s Z s ^ '&y dddd / :dd de dede dedd ^ > d / W : ^ Z W < > d ddde / ' : ^ dde d dee ddd Page 20
21 ^ > d ^ Z > d / ' D :, d : W < ddde W E ^ h^ddd dd deddd dedde ^ ^, D D & ^ d W <, <W dddd > ' d : dee de ddeee ddedd ^ W W :,, Wt < z: < E,t, <W KW ddde ' E ddd eddd dee ddd ^ '&y dddd ^ / K ^ dd d dde ddd ^ ^, Z ddde / ' d dd dd dd ded ded ^ : y ' W D K ^ '& ddde : ded d ddee ddee ^ Z ddde ed W e edd edd d K d W < ^ Z dddd ' 'WZ dd de eeed eeed d z d d ^ ^ z deee ^ ' &^> dde d d ddd ddd d ^ D dddd Z ' DKZ d d dd dd d ', ' D D Z W < ddde Z W d de dee ddd s : D D t dddd d ^ ded ddde dddd dddd t d ^ s D: :' D Z W, Z > 't d ' ^ '&y ddde ^ d ' E ddd eddd dee ded t ' < Z ^ dddd d ' d e d ddd dde t ^<& Z D deed D Z d & W ' W : dee de deeee deeee z, < D D < d K deed dd de ddeed ddeee Page 21
22 Footnotes: This research was supported by NIH grants EY09339, GM , and by an unrestricted grant from Amgen Inc and Sandler Program for Asthma Research. DM was supported in part by the CWRU MSTP and NIH T32 GM Page 22
23 Legends for Figures Figure 1. Structural transformations of rhodopsin during photolysis. Alignment of the first three photointermediates, bovine rhodopsin colored by helix: helix I (blue), helix II (cyan), helix III (violet), helix IV (red), helix V (orange), helix VI (yellow), helix VII (green), and helix 8 (magenta); bovine bathorhodopsin (magenta); and bovine lumirhodopsin (green), reveals that the most pronounced structural changes occur in the chromophore region (dashed rectangle). The chromophore from each of the three structures is shown on the right to highlight the changes as photolysis proceeds. Retinal is colored red for ground state rhodopsin, magenta for bathorhodopsin, and green for lumirhodopsin. From the ground state to bathorhodopsin, the C 11 =C 12 bond adopts a trans configuration after illumination. From bathorhodopsin to lumirhodopsin, a conformational change in the β-ionone ring of the retinal is apparent. Changes in meta II and opsin are more pronounced, with opsin showing slight side chain shifts of helices surrounding the chromophore binding site that allow retinal entry and exit as illustrated in Figure 4. Figure 2. Comparison of rhodopsin with the photoactivated state resembling the meta II structure and squid rhodopsin. (A) Side view. Molecules are colored as the bovine photoactivated structure (green ribbon) and bovine rhodopsin by helix I (blue), helix II (cyan), helix III (violet), helix IV (red), helix V (orange), helix VI (yellow), helix VII (green), and helix 8 (magenta). These structures revealed smaller conformational changes than predicted from EPR studies. (B) Cytoplasmic view. Although the photoactivated structure resembling meta II is structurally similar to that of the ground-state rhodopsin, portions of the C-II and C-III loops are disordered in the photoactive state. The C-III loop does not even follow the path of the loop found in the ground-state crystals. This difference may reflect the importance of these loops dynamics in transducin activation. (C) Side view of squid rhodopsin is shown as a green ribbon overlying bovine rhodopsin colored by helix. (D) These two structures show immense structural homology, with the most noticeable difference being the larger C-III loop in squid rhodopsin due to an extra sequence in that region. The difference in the C-III loop, involved in the binding of transducin, suggests that this may be an important structural motif for specifying different modes of coupling with G q -type G proteins in invertebrate rhodopsins. Page 23
24 Figure 3. Comparison of rhodopsin and opsin structures. (A) Side view. Molecules colored as bovine opsin II (green ribbon) and bovine rhodopsin by helix: helix I (blue), helix II (cyan), helix III (violet), helix IV (red), helix V (orange), helix VI (yellow), helix VII (green), and helix 8 (magenta). Despite overall helical plasticity, the structural overlay illustrates slight shifts in the side chains that could explain retinal entry/exit (see Figure 4). (B) Cytoplasmic view. The opsin and rhodopsin structures show similar tracings of the cytoplasmic loops, but slight differences may indicate the importance of these loop movements in activation or relaxation back to the ground state. Figure 4. Structural comparison between opsin and rhodopsin illustrating a pathway for retinal exchange. The opsin structure shows two different openings to the retinal-binding pocket, i.e. one between the extracellular ends of helix V and helix VI and the other between helix I and helix VII. These openings suggest different chromophore entrance and exit routes. Both bovine opsin and bovine rhodopsin are colored by helix: helix I (blue), helix II (cyan), helix III (violet), helix IV (red), helix V (orange), helix VI (yellow), helix VII (green), and helix 8 (magenta), while retinal is depicted as red sticks. (A) Use of the online software Caver shows a tunneling path of retinal through an entry site between helix V and helix VI of opsin, with three key Phe residues lining the path (Phe 208, Phe 212, and Phe 273 ). (B) This retinal entry path is blocked in rhodopsin due to shifts in helix V and helix VI that cause repositioning of these three key Phe residues. (C) A distinct pathway for retinal exit from opsin is formed between helix I and helix VII and is illustrated as a clear hole in the solvent accessible surface. (D) Similar to the retinal entry pathway, the exit pathway is closed off in rhodopsin, most notably by a shift in Phe 293, as evidenced by the closed solvent accessible surface. Figure 5. Comparison of β 1 -adrenergic receptor with rhodopsin, β 2 -adrenergic receptor, and A 2A adenosine receptor structures. The turkey β 1 -adrenergic receptor was mutated to facilitate its crystallization. The mutated receptor evidenced enhanced thermostability and preferentially existed in an antagonist-binding state. Stretches of amino acid sequence were deleted from the Page 24
25 N-terminal region, i.e. the loop connecting helix V and helix VI, and the C-terminal region. (A) The β 1 -adrenergic receptor is colored by helix: helix I (blue), helix II (cyan), helix III (violet), helix IV (red), helix V (orange), helix VI (yellow), helix VII (green), and helix VIII (magenta). Thermostabilizing mutations (R68S, M90V, Y227A, A282L (not resolved in the shown structure), F327A, and F338M) and those that either increased functional expression (C116L) or eliminated a palmitoylation site (C358A) are shown as balls/sticks. (B) Structure of the β 1 - adrenergic receptor (colored by helix) is shown with bovine rhodopsin (green ribbon) to illustrate key structural differences. Despite some differences in the transmembrane helices, the main distinction is in the organization of the cytoplasmic (C-) loops. The C-II loop in the β 1 -adrenergic receptor structure forms a short α-helix whereas this loop is more extended in rhodopsin. Also, rhodopsin has a native C-III loop that is absent from the β 1 -adrenergic receptor structure due to deletions needed for crystallization. (C) The structure of the β 1 -adrenergic receptor (colored by helix) is displayed with the human β2-adrenergic receptor (green ribbon) to show the key structural differences. In this case, the C-II loops of both the β 1 -adrenergic receptor and β 2 - adrenergic receptor are similar, but the α-helical conformation of the β 1 -adrenergic receptor cannot be accommodated within the two crystallized structures of the β 2 -adrenergic receptor because of lattice contacts with adjacent molecules. (D) The structure of the β 1 -adrenergic receptor (colored by helix) is displayed with the human A 2A adenosine receptor (green ribbon) to show key structural differences. The main difference between these two structures lies in the distinct folding of the extracellular loops, with the adenosine receptor adopting a more open surface for ligand binding due to this folding. Figure 6. Structural comparison of the β 2 -adrenergic and A 2A adenosine fusion receptors with rhodopsin. Shown are structures of the human β 2 -adrenergic receptor fused to T4 lysozyme (A) and in complex with Fab (B) and the human A 2A adenosine receptor fused to T4 lysozyme (C). Bovine rhodopsin is colored by helix: helix I (blue), helix II (cyan), helix III (violet), helix IV (red), helix V (orange), helix VI (yellow), helix VII (green), and helix 8 (magenta) whereas each of the two fusion β 2 -adrenergic receptors are displayed as green ribbons. There are differences in the transmembrane helices between the β 2 -adrenergic receptor and rhodopsin, with the most pronounced disparity occurring in helix I because the Pro residue kink is absent in the β 2 - adrenergic receptor structures. Also, the β 2 -adrenergic receptor structures show minimal inter- Page 25
26 receptor contacts illustrating that protein engineering may have affected the crystal packing and disrupted any possible dimeric interfaces of these structures. The overall helical differences between the adenosine receptor and rhodopsin are minimal, but the adenosine receptor has a distinct arrangement of the extracellular loops for a more open structure of the ligand binding pocket that is shifted closer to helices VI and VII. Page 26
27 Table 1. Characteristics of class A G protein-coupled-receptor crystal structures. Structure Release Year PDB ID Resolution/Space Group Mutations Truncations Unresolved Residues Ligands/complexes or fusions/other alternations bovine rhodopsin f Ǻ /P Q236-E239, L328-A cis-retinal bovine rhodopsin a gzm 2.65 Ǻ /P P327-G329, A333-A cis-retinal bovine rhodopsin u Ǻ /P cis-retinal 9-cis-bovine rhodopsin (isorhodopsin) recombinant bovine rhodopsin mutant ped 2.95 Ǻ /P cis-retinal j4y 3.40 Ǻ /P3 1 N2C, D282C - L328-A cis-retinal squid rhodopsin z Ǻ /P6 2 - S359-A448 M1-N8 11-cis-retinal squid rhodopsin ziy 3.70 Ǻ /C V18I M374-A448 G2-R3 11-cis-retinal bovine bathorhodopsin bovine lumirhodoposin bovine meta II-like rhodopsin (photoactivated) g Ǻ /P all-trans-retinal hpy 2.80 Ǻ /P all-trans-retinal i Ǻ /P V230-Q238, K311-F313, D330-A348 all-trans-retinal opsin cap 2.90 Ǻ /H3 - - P327-A348 - Molecular Pharmacology Fast Forward. Published on October 22, 2008 as DOI: /mol Page 27
28 opsin dqb 3.20 Ǻ /H 3 2 K341L - P327-A348 C-terminal peptide (11 mer) derived from the Galpha subunit of transducin turkey β 1 -adrenergic receptor human β 2 - adrenergic receptor human β 2 - adrenergic receptor human β 2 - adrenergic receptor vt Ǻ/P1 R68S, M90V, C116L, Y227A, A282L, F327A, F338M, C358A, and deglycosylated Page 28 D2-S32, C244-R271, A368-A496 M1-G2, A33-Q39, R239-R243, A272- M283, P360-L r4r 3.40 Ǻ /C 2 (C 121) - Y366-L413 M1-M36, K60, A91- E107, S165-A202, Q243-K263, V292- L310, A349-G rh Ǻ / C 2 (C 121) N187E (elimination of glycosylation site), Q231-S262 (replaced by amino acids 2-161of T4 lysozyme), C1054T and C1097A (lysozyme) d4s 2.80 Ǻ /P E122W, N187E (elimination of glycosylation site), Q231-S262 (replaced by amino acids 2-161of T4 lysozyme),c1054t and C1097A (lysozyme) cyanopindolol carazolol; Fab attachment to C-III loop and portion of helix VII Y366-L413 M1-R28, R343-G365 carazolol; T4 lysozyme chimera to C-III loop R349-L413 M1-V31, R343-K348 timolol; T4 lysozyme chimera to C-III loop Molecular Pharmacology Fast Forward. Published on October 22, 2008 as DOI: /mol
29 ZM antagonist; T4 lysozyme chimera to C-III loop Molecular Pharmacology Fast Forward. Published on October 22, 2008 as DOI: /mol a human A2A adenoside receptor eml 2.60 Ǻ /P L209-A221 (replaced by amino acids of T4 lysozyme), C1054T and C1097A (lysozyme) A317-S412 M1-P2, Q148-S156, Q311-A316 Corrected space group by (Stenkamp, 2008). Page 29

30 Rhodopsin: 11-cis-retinylidene Batho: all-trans-retinylidene Lumi: all-trans-retinylidene Figure 1


 C D C-III")

31 A B C-III C-II Figure 2 bovine rhodopsin (rainbow helices) photoactivated structure (green ribbon) C D C-III bovine rhodopsin (rainbow helices) squid rhodopsin (green ribbon)
 bovine opsin (green")
32 Figure 3 A B bovine rhodopsin (rainbow helices) bovine opsin (green ribbon)



33 A opsin rhodopsin B Figure 4 Phe 212 Phe 208 Phe 212 Phe 208 Phe 273 Phe 273 C opsin D rhodopsin Tyr 43 Phe 293 Tyr 43 Phe 293 Leu 40 Leu 40




34 Figure 5 B C A Leu116 Ala327 Val90 rhodopsin (green ribbon) D β 2 -adrenergic receptor (green ribbon) Ala227 Met338 Ser68 Ala358 β 1 -adrenergic receptor (rainbow helices) adenosine A 2A receptor (green ribbon)
 β 2 -adrenergic")
 (B)")

35 Figure 6 A B C bovine rhodopsin (rainbow helices) β 2 -adrenergic receptors fused with T4-lysozyme (A) or with Fab (green ribbons) (B) bovine rhodopsin (rainbow helices) Αdenosine A 2A receptor fused with T4-lysozyme (C)
Topology of Class A G Protein-Coupled Receptors: Insights Gained from Crystal Structures of Rhodopsins, Adrenergic and Adenosine Receptors
 0026-895X/09/7501-1 12$20.00 MOLECULAR PHARMACOLOGY Vol. 75, No. 1 Copyright 2009 The American Society for Pharmacology and Experimental Therapeutics 51938/3423246 Mol Pharmacol 75:1 12, 2009 Printed in
0026-895X/09/7501-1 12$20.00 MOLECULAR PHARMACOLOGY Vol. 75, No. 1 Copyright 2009 The American Society for Pharmacology and Experimental Therapeutics 51938/3423246 Mol Pharmacol 75:1 12, 2009 Printed in
Visual pigments. Neuroscience, Biochemistry Dr. Mamoun Ahram Third year, 2019
 Visual pigments Neuroscience, Biochemistry Dr. Mamoun Ahram Third year, 2019 References Webvision: The Organization of the Retina and Visual System (http://www.ncbi.nlm.nih.gov/books/nbk11522/#a 127) The
Visual pigments Neuroscience, Biochemistry Dr. Mamoun Ahram Third year, 2019 References Webvision: The Organization of the Retina and Visual System (http://www.ncbi.nlm.nih.gov/books/nbk11522/#a 127) The
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11085 Supplementary Tables: Supplementary Table 1. Summary of crystallographic and structure refinement data Structure BRIL-NOP receptor Data collection Number of crystals 23 Space group
doi:10.1038/nature11085 Supplementary Tables: Supplementary Table 1. Summary of crystallographic and structure refinement data Structure BRIL-NOP receptor Data collection Number of crystals 23 Space group
Visual pigments. Neuroscience, Biochemistry Dr. Mamoun Ahram Third year, 2015
 Visual pigments Neuroscience, Biochemistry Dr. Mamoun Ahram Third year, 2015 References Photoreceptors and visual pigments Webvision: The Organization of the Retina and Visual System (http://www.ncbi.nlm.nih.gov/books/nbk11522/#a127)
Visual pigments Neuroscience, Biochemistry Dr. Mamoun Ahram Third year, 2015 References Photoreceptors and visual pigments Webvision: The Organization of the Retina and Visual System (http://www.ncbi.nlm.nih.gov/books/nbk11522/#a127)
Nature Structural & Molecular Biology: doi: /nsmb Supplementary Figure 1
 Supplementary Figure 1 Crystallization. a, Crystallization constructs of the ET B receptor are shown, with all of the modifications to the human wild-type the ET B receptor indicated. Residues interacting
Supplementary Figure 1 Crystallization. a, Crystallization constructs of the ET B receptor are shown, with all of the modifications to the human wild-type the ET B receptor indicated. Residues interacting
Protein Structure. W. M. Grogan, Ph.D. OBJECTIVES
 Protein Structure W. M. Grogan, Ph.D. OBJECTIVES 1. Describe the structure and characteristic properties of typical proteins. 2. List and describe the four levels of structure found in proteins. 3. Relate
Protein Structure W. M. Grogan, Ph.D. OBJECTIVES 1. Describe the structure and characteristic properties of typical proteins. 2. List and describe the four levels of structure found in proteins. 3. Relate
Computational modeling of G-Protein Coupled Receptors (GPCRs) has recently become
 has recently become") Homology Modeling and Docking of Melatonin Receptors Andrew Kohlway, UMBC Jeffry D. Madura, Duquesne University 6/18/04 INTRODUCTION Computational modeling of G-Protein Coupled Receptors (GPCRs) has recently
Homology Modeling and Docking of Melatonin Receptors Andrew Kohlway, UMBC Jeffry D. Madura, Duquesne University 6/18/04 INTRODUCTION Computational modeling of G-Protein Coupled Receptors (GPCRs) has recently
Bahnson Biochemistry Cume, April 8, 2006 The Structural Biology of Signal Transduction
 Name page 1 of 6 Bahnson Biochemistry Cume, April 8, 2006 The Structural Biology of Signal Transduction Part I. The ion Ca 2+ can function as a 2 nd messenger. Pick a specific signal transduction pathway
Name page 1 of 6 Bahnson Biochemistry Cume, April 8, 2006 The Structural Biology of Signal Transduction Part I. The ion Ca 2+ can function as a 2 nd messenger. Pick a specific signal transduction pathway
β1 Structure Prediction and Validation
 13 Chapter 2 β1 Structure Prediction and Validation 2.1 Overview Over several years, GPCR prediction methods in the Goddard lab have evolved to keep pace with the changing field of GPCR structure. Despite
13 Chapter 2 β1 Structure Prediction and Validation 2.1 Overview Over several years, GPCR prediction methods in the Goddard lab have evolved to keep pace with the changing field of GPCR structure. Despite
Activation of G Protein Coupled Receptors: Beyond Two-State Models and Tertiary Conformational Changes
 Annu. Rev. Pharmacol. Toxicol. 2008. 48:107 41 First published online as a Review in Advance on September 10, 2007 The Annual Review of Pharmacology and Toxicology is online at http://pharmtox.annualreviews.org
Annu. Rev. Pharmacol. Toxicol. 2008. 48:107 41 First published online as a Review in Advance on September 10, 2007 The Annual Review of Pharmacology and Toxicology is online at http://pharmtox.annualreviews.org
Signal Transduction Phosphorylation Protein kinases. Misfolding diseases. Protein Engineering Lysozyme variants
 Signal Transduction Phosphorylation Protein kinases Misfolding diseases Protein Engineering Lysozyme variants Cells and Signals Regulation The cell must be able to respond to stimuli Cellular activities
Signal Transduction Phosphorylation Protein kinases Misfolding diseases Protein Engineering Lysozyme variants Cells and Signals Regulation The cell must be able to respond to stimuli Cellular activities
MOLECULAR DRUG TARGETS
 MOLECULAR DRUG TARGETS LEARNING OUTCOMES At the end of this session student shall be able to: List different types of druggable targets Describe forces involved in drug-receptor interactions Describe theories
MOLECULAR DRUG TARGETS LEARNING OUTCOMES At the end of this session student shall be able to: List different types of druggable targets Describe forces involved in drug-receptor interactions Describe theories
Comparison between Bacteriorhodopsin and Halorhodopsin. Halorhodopsin (HR) and Bacteriorhodopsin (BR) belong to a subfamily of
 and Bacteriorhodopsin (BR) belong to a subfamily of") Comparison between Bacteriorhodopsin and Halorhodopsin Halorhodopsin (HR) and Bacteriorhodopsin (BR) belong to a subfamily of heptahelical membrane proteins, the archaeal rhodopsins. They are found in
Comparison between Bacteriorhodopsin and Halorhodopsin Halorhodopsin (HR) and Bacteriorhodopsin (BR) belong to a subfamily of heptahelical membrane proteins, the archaeal rhodopsins. They are found in
RHODOPSIN: Insights from Recent Structural Studies
 Annu. Rev. Biophys. Biomol. Struct. 2002. 31:443 84 DOI: 10.1146/annurev.biophys.31.082901.134348 Copyright c 2002 by Annual Reviews. All rights reserved RHODOPSIN: Insights from Recent Structural Studies
Annu. Rev. Biophys. Biomol. Struct. 2002. 31:443 84 DOI: 10.1146/annurev.biophys.31.082901.134348 Copyright c 2002 by Annual Reviews. All rights reserved RHODOPSIN: Insights from Recent Structural Studies
h D t del d ddl ddl el del d < d
 > Z Z d : d h D d, Z d d / d W < ^ d d d ' e W E e > ^ d d / & d e ^ < e Z d e & D d e h s d : ^ d e d ' D ' h ' ' ' d / / ' h ^ ^ ' d / / ' & ' h D ' ' ' d W ' h ' ' ' d ' D ' ' > ' h ' ' ' e D /' /EZh
> Z Z d : d h D d, Z d d / d W < ^ d d d ' e W E e > ^ d d / & d e ^ < e Z d e & D d e h s d : ^ d e d ' D ' h ' ' ' d / / ' h ^ ^ ' d / / ' & ' h D ' ' ' d W ' h ' ' ' d ' D ' ' > ' h ' ' ' e D /' /EZh
Model Worksheet Teacher Key
 Introduction Despite the complexity of life on Earth, the most important large molecules found in all living things (biomolecules) can be classified into only four main categories: carbohydrates, lipids,
Introduction Despite the complexity of life on Earth, the most important large molecules found in all living things (biomolecules) can be classified into only four main categories: carbohydrates, lipids,
New G-protein-coupled receptor crystal structures: insights and limitations
 Opinion New G-protein-coupled receptor crystal structures: insights and limitations Brian Kobilka 1 and Gebhard F.X. Schertler 2 1 Departments of Molecular and Cellular Physiology and Medicine, Stanford
Opinion New G-protein-coupled receptor crystal structures: insights and limitations Brian Kobilka 1 and Gebhard F.X. Schertler 2 1 Departments of Molecular and Cellular Physiology and Medicine, Stanford
Biotechnology of Proteins. The Source of Stability in Proteins (III) Fall 2015
 Fall 2015") Biotechnology of Proteins The Source of Stability in Proteins (III) Fall 2015 Conformational Entropy of Unfolding It is The factor that makes the greatest contribution to stabilization of the unfolded
Biotechnology of Proteins The Source of Stability in Proteins (III) Fall 2015 Conformational Entropy of Unfolding It is The factor that makes the greatest contribution to stabilization of the unfolded
SUPPLEMENTARY INFORMATION
 SUPPLEMENTARY INFORMATION doi:10.1038/nature11524 Supplementary discussion Functional analysis of the sugar porter family (SP) signature motifs. As seen in Fig. 5c, single point mutation of the conserved
SUPPLEMENTARY INFORMATION doi:10.1038/nature11524 Supplementary discussion Functional analysis of the sugar porter family (SP) signature motifs. As seen in Fig. 5c, single point mutation of the conserved
Retinal Proteins (Rhodopsins) Vision, Bioenergetics, Phototaxis. Bacteriorhodopsin s Photocycle. Bacteriorhodopsin s Photocycle
 Vision, Bioenergetics, Phototaxis. Bacteriorhodopsin s Photocycle. Bacteriorhodopsin s Photocycle") Molecular chanisms of Photoactivation and Spectral Tuning in Retinal Proteins Emad Tajkhorshid Theoretical and Computational Biophysics Group Beckman Institute University of Illinois at Urbana-Champaign
Molecular chanisms of Photoactivation and Spectral Tuning in Retinal Proteins Emad Tajkhorshid Theoretical and Computational Biophysics Group Beckman Institute University of Illinois at Urbana-Champaign
An Introduction to Metabolism
 An Introduction to Metabolism I. All of an organism=s chemical reactions taken together is called metabolism. A. Metabolic pathways begin with a specific molecule, which is then altered in a series of
An Introduction to Metabolism I. All of an organism=s chemical reactions taken together is called metabolism. A. Metabolic pathways begin with a specific molecule, which is then altered in a series of
Supplementary information for:
 SUPPLEMETARY IFRMATI Supplementary information for: Structure of a β 1 -adrenergic G protein-coupled receptor Tony Warne, Maria J. Serrano-Vega, Jillian G. Baker#, Rouslan Moukhametzianov, Patricia C.
SUPPLEMETARY IFRMATI Supplementary information for: Structure of a β 1 -adrenergic G protein-coupled receptor Tony Warne, Maria J. Serrano-Vega, Jillian G. Baker#, Rouslan Moukhametzianov, Patricia C.
Computational Studies of the Photoreceptor Rhodopsin. Scott E. Feller Wabash College
 Computational Studies of the Photoreceptor Rhodopsin Scott E. Feller Wabash College Rhodopsin Photocycle Dark-adapted Rhodopsin hn Isomerize retinal Photorhodopsin ~200 fs Bathorhodopsin Meta-II ms timescale
Computational Studies of the Photoreceptor Rhodopsin Scott E. Feller Wabash College Rhodopsin Photocycle Dark-adapted Rhodopsin hn Isomerize retinal Photorhodopsin ~200 fs Bathorhodopsin Meta-II ms timescale
Supplementary Figure S1. Urea-mediated buffering mechanism of H. pylori. Gastric urea is funneled to a cytoplasmic urease that is presumably attached
 Supplementary Figure S1. Urea-mediated buffering mechanism of H. pylori. Gastric urea is funneled to a cytoplasmic urease that is presumably attached to HpUreI. Urea hydrolysis products 2NH 3 and 1CO 2
Supplementary Figure S1. Urea-mediated buffering mechanism of H. pylori. Gastric urea is funneled to a cytoplasmic urease that is presumably attached to HpUreI. Urea hydrolysis products 2NH 3 and 1CO 2
Transmembrane Domains (TMDs) of ABC transporters
 of ABC transporters") Transmembrane Domains (TMDs) of ABC transporters Most ABC transporters contain heterodimeric TMDs (e.g. HisMQ, MalFG) TMDs show only limited sequence homology (high diversity) High degree of conservation
Transmembrane Domains (TMDs) of ABC transporters Most ABC transporters contain heterodimeric TMDs (e.g. HisMQ, MalFG) TMDs show only limited sequence homology (high diversity) High degree of conservation
Potassium channel gating and structure!
 Reading: Potassium channel gating and structure Hille (3rd ed.) chapts 10, 13, 17 Doyle et al. The Structure of the Potassium Channel: Molecular Basis of K1 Conduction and Selectivity. Science 280:70-77
Reading: Potassium channel gating and structure Hille (3rd ed.) chapts 10, 13, 17 Doyle et al. The Structure of the Potassium Channel: Molecular Basis of K1 Conduction and Selectivity. Science 280:70-77
2. In regards to the fluid mosaic model, which of the following is TRUE?
 General Biology: Exam I Sample Questions 1. How many electrons are required to fill the valence shell of a neutral atom with an atomic number of 24? a. 0 the atom is inert b. 1 c. 2 d. 4 e. 6 2. In regards
General Biology: Exam I Sample Questions 1. How many electrons are required to fill the valence shell of a neutral atom with an atomic number of 24? a. 0 the atom is inert b. 1 c. 2 d. 4 e. 6 2. In regards
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature17991 Supplementary Discussion Structural comparison with E. coli EmrE The DMT superfamily includes a wide variety of transporters with 4-10 TM segments 1. Since the subfamilies of the
doi:10.1038/nature17991 Supplementary Discussion Structural comparison with E. coli EmrE The DMT superfamily includes a wide variety of transporters with 4-10 TM segments 1. Since the subfamilies of the
Biophysics 490M Project
 Biophysics 490M Project Dan Han Department of Biochemistry Structure Exploration of aa 3 -type Cytochrome c Oxidase from Rhodobacter sphaeroides I. Introduction: All organisms need energy to live. They
Biophysics 490M Project Dan Han Department of Biochemistry Structure Exploration of aa 3 -type Cytochrome c Oxidase from Rhodobacter sphaeroides I. Introduction: All organisms need energy to live. They
Copyright Mark Brandt, Ph.D A third method, cryogenic electron microscopy has seen increasing use over the past few years.
 Structure Determination and Sequence Analysis The vast majority of the experimentally determined three-dimensional protein structures have been solved by one of two methods: X-ray diffraction and Nuclear
Structure Determination and Sequence Analysis The vast majority of the experimentally determined three-dimensional protein structures have been solved by one of two methods: X-ray diffraction and Nuclear
SUPPLEMENTARY INFORMATION
 Supplementary Results DNA binding property of the SRA domain was examined by an electrophoresis mobility shift assay (EMSA) using synthesized 12-bp oligonucleotide duplexes containing unmodified, hemi-methylated,
Supplementary Results DNA binding property of the SRA domain was examined by an electrophoresis mobility shift assay (EMSA) using synthesized 12-bp oligonucleotide duplexes containing unmodified, hemi-methylated,
Minireview: Molecular Structure and Dynamics of Drug Targets
 Prague Medical Report / Vol. 109 (2008) No. 2 3, p. 107 112 107) Minireview: Molecular Structure and Dynamics of Drug Targets Dahl S. G., Sylte I. Department of Pharmacology, Institute of Medical Biology,
Prague Medical Report / Vol. 109 (2008) No. 2 3, p. 107 112 107) Minireview: Molecular Structure and Dynamics of Drug Targets Dahl S. G., Sylte I. Department of Pharmacology, Institute of Medical Biology,
Regulation and signaling. Overview. Control of gene expression. Cells need to regulate the amounts of different proteins they express, depending on
 Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Regulation and signaling Overview Cells need to regulate the amounts of different proteins they express, depending on cell development (skin vs liver cell) cell stage environmental conditions (food, temperature,
Heterotrimeric G proteins and the role of lipids in signaling. John Sondek, Ph.D. Depts. of Pharmacology and Biochemistry & Biophyscis
 Heterotrimeric G proteins and the role of lipids in signaling John Sondek, Ph.D. Depts. of Pharmacology and Biochemistry & Biophyscis The GTPase cycle molecular switch A GTPases is NOT a kinase Two major
Heterotrimeric G proteins and the role of lipids in signaling John Sondek, Ph.D. Depts. of Pharmacology and Biochemistry & Biophyscis The GTPase cycle molecular switch A GTPases is NOT a kinase Two major
Molecular Pharmacology Fast Forward. Published on June 29, 2016 as DOI: /mol This article has not been copyedited and formatted.
 , W D E / & h E D D D Z W, W D / & h dddd h D D d dddddedddede dddddeddddee & dddddedddede d d d & d dee d eddd Z dde DW d d d W/ d d e e d d d d d, d /^ d db d db E< E l
, W D E / & h E D D D Z W, W D / & h dddd h D D d dddddedddede dddddeddddee & dddddedddede d d d & d dee d eddd Z dde DW d d d W/ d d e e d d d d d, d /^ d db d db E< E l
Introduction to Comparative Protein Modeling. Chapter 4 Part I
 Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
Introduction to Comparative Protein Modeling Chapter 4 Part I 1 Information on Proteins Each modeling study depends on the quality of the known experimental data. Basis of the model Search in the literature
MOLECULAR DYNAMICS SIMULATIONS ON STRUCTURAL CONFORMATIONS OF RHODOPSIN AND PRION PROTEINS
 Ó³ Ÿ. 2005.. 2, º 6(129).. 87Ä97 Š [51-72:530.145]+[51-72:541.1] MOLECULAR DYNAMICS SIMULATIONS ON STRUCTURAL CONFORMATIONS OF RHODOPSIN AND PRION PROTEINS Kh. T. Kholmurodov 1 Joint Institute for Nuclear
Ó³ Ÿ. 2005.. 2, º 6(129).. 87Ä97 Š [51-72:530.145]+[51-72:541.1] MOLECULAR DYNAMICS SIMULATIONS ON STRUCTURAL CONFORMATIONS OF RHODOPSIN AND PRION PROTEINS Kh. T. Kholmurodov 1 Joint Institute for Nuclear
ml. ph 7.5 ph 6.5 ph 5.5 ph 4.5. β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS
 a UV28 absorption (mau) 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS β 2 AR-Gs complex dissociated complex excess nucleotides b 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex
a UV28 absorption (mau) 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex + GDP β 2 AR-Gs complex + GTPγS β 2 AR-Gs complex dissociated complex excess nucleotides b 9 8 7 5 3 β 2 AR-Gs complex β 2 AR-Gs complex
Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA
 Online Supplemental Results Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA Structure solution and overall architecture
Online Supplemental Results Detailed description of overall and active site architecture of PPDC- 3dThDP, PPDC-2HE3dThDP, PPDC-3dThDP-PPA and PPDC- 3dThDP-POVA Structure solution and overall architecture
Patrick: An Introduction to Medicinal Chemistry 5e Chapter 04
 01) Which of the following statements is not true about receptors? a. Most receptors are proteins situated inside the cell. b. Receptors contain a hollow or cleft on their surface which is known as a binding
01) Which of the following statements is not true about receptors? a. Most receptors are proteins situated inside the cell. b. Receptors contain a hollow or cleft on their surface which is known as a binding
Chapter 6- An Introduction to Metabolism*
 Chapter 6- An Introduction to Metabolism* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. The Energy of Life
Chapter 6- An Introduction to Metabolism* *Lecture notes are to be used as a study guide only and do not represent the comprehensive information you will need to know for the exams. The Energy of Life
1. What is an ångstrom unit, and why is it used to describe molecular structures?
 1. What is an ångstrom unit, and why is it used to describe molecular structures? The ångstrom unit is a unit of distance suitable for measuring atomic scale objects. 1 ångstrom (Å) = 1 10-10 m. The diameter
1. What is an ångstrom unit, and why is it used to describe molecular structures? The ångstrom unit is a unit of distance suitable for measuring atomic scale objects. 1 ångstrom (Å) = 1 10-10 m. The diameter
GPCR Dynamics: Structures in Motion
 This is an open access article published under an ACS AuthorChoice License, which permits copying and redistribution of the article or any adaptations for non-commercial purposes. pubs.acs.org/cr GPCR
This is an open access article published under an ACS AuthorChoice License, which permits copying and redistribution of the article or any adaptations for non-commercial purposes. pubs.acs.org/cr GPCR
LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor
 LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor Note: Adequate space is given for each answer. Questions that require a brief explanation should
LS1a Fall 2014 Problem Set #2 Due Monday 10/6 at 6 pm in the drop boxes on the Science Center 2 nd Floor Note: Adequate space is given for each answer. Questions that require a brief explanation should
Examples of Protein Modeling. Protein Modeling. Primary Structure. Protein Structure Description. Protein Sequence Sources. Importing Sequences to MOE
 Examples of Protein Modeling Protein Modeling Visualization Examination of an experimental structure to gain insight about a research question Dynamics To examine the dynamics of protein structures To
Examples of Protein Modeling Protein Modeling Visualization Examination of an experimental structure to gain insight about a research question Dynamics To examine the dynamics of protein structures To
Section B. Multi-Pass Receptors. James A. Wells, Editor BRADSHAW
 c0002 Section B Multi-Pass Receptors p0010 James A. Wells, Editor BRADSHAW 978-0-12-374145-5 00388 SEC2.indd 149 4/22/2009 3:06:25 PM SEC2.indd 150 4/22/2009 3:06:25 PM Chapter 23 c00023 Structure and
c0002 Section B Multi-Pass Receptors p0010 James A. Wells, Editor BRADSHAW 978-0-12-374145-5 00388 SEC2.indd 149 4/22/2009 3:06:25 PM SEC2.indd 150 4/22/2009 3:06:25 PM Chapter 23 c00023 Structure and
Table 1. Kinetic data obtained from SPR analysis of domain 11 mutants interacting with IGF-II. Kinetic parameters K D 1.
 Kinetics and Thermodynamics of the Insulin-like Growth Factor II (IGF-II) Interaction with IGF-II/Mannose 6-phosphate Receptor and the function of CD and AB Loop Solvent-exposed Residues. Research Team:
Kinetics and Thermodynamics of the Insulin-like Growth Factor II (IGF-II) Interaction with IGF-II/Mannose 6-phosphate Receptor and the function of CD and AB Loop Solvent-exposed Residues. Research Team:
Solutions and Non-Covalent Binding Forces
 Chapter 3 Solutions and Non-Covalent Binding Forces 3.1 Solvent and solution properties Molecules stick together using the following forces: dipole-dipole, dipole-induced dipole, hydrogen bond, van der
Chapter 3 Solutions and Non-Covalent Binding Forces 3.1 Solvent and solution properties Molecules stick together using the following forces: dipole-dipole, dipole-induced dipole, hydrogen bond, van der
9/25/2011. Outline. Overview: The Energy of Life. I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V.
 Chapter 8 Introduction to Metabolism Outline I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V. Enzymes Overview: The Energy of Life Figure 8.1 The living cell is a miniature
Chapter 8 Introduction to Metabolism Outline I. Forms of Energy II. Laws of Thermodynamics III. Energy and metabolism IV. ATP V. Enzymes Overview: The Energy of Life Figure 8.1 The living cell is a miniature
BIOCHEMISTRY GUIDED NOTES - AP BIOLOGY-
 BIOCHEMISTRY GUIDED NOTES - AP BIOLOGY- ELEMENTS AND COMPOUNDS - anything that has mass and takes up space. - cannot be broken down to other substances. - substance containing two or more different elements
BIOCHEMISTRY GUIDED NOTES - AP BIOLOGY- ELEMENTS AND COMPOUNDS - anything that has mass and takes up space. - cannot be broken down to other substances. - substance containing two or more different elements
Quantitative Stability/Flexibility Relationships; Donald J. Jacobs, University of North Carolina at Charlotte Page 1 of 12
 Quantitative Stability/Flexibility Relationships; Donald J. Jacobs, University of North Carolina at Charlotte Page 1 of 12 The figure shows that the DCM when applied to the helix-coil transition, and solved
Quantitative Stability/Flexibility Relationships; Donald J. Jacobs, University of North Carolina at Charlotte Page 1 of 12 The figure shows that the DCM when applied to the helix-coil transition, and solved
Ligand screening system using fusion proteins of G protein coupled receptors with G protein α subunits
 2 Ligand screening system using fusion proteins of G protein coupled receptors with G protein α subunits G protein coupled receptors A key player of signaling transduction. Cell membranes are packed with
2 Ligand screening system using fusion proteins of G protein coupled receptors with G protein α subunits G protein coupled receptors A key player of signaling transduction. Cell membranes are packed with
Homology Models of Human Adrenergic Receptors
 58 Chapter 4 Homology Models of Human Adrenergic Receptors The increased availability of high-resolution GPCR crystal structures has enabled deeper investigation into homology modeling as a method for
58 Chapter 4 Homology Models of Human Adrenergic Receptors The increased availability of high-resolution GPCR crystal structures has enabled deeper investigation into homology modeling as a method for
Supplementary information
 Supplementary information The structural basis of modularity in ECF-type ABC transporters Guus B. Erkens 1,2, Ronnie P-A. Berntsson 1,2, Faizah Fulyani 1,2, Maria Majsnerowska 1,2, Andreja Vujičić-Žagar
Supplementary information The structural basis of modularity in ECF-type ABC transporters Guus B. Erkens 1,2, Ronnie P-A. Berntsson 1,2, Faizah Fulyani 1,2, Maria Majsnerowska 1,2, Andreja Vujičić-Žagar
Receptor Based Drug Design (1)
") Induced Fit Model For more than 100 years, the behaviour of enzymes had been explained by the "lock-and-key" mechanism developed by pioneering German chemist Emil Fischer. Fischer thought that the chemicals
Induced Fit Model For more than 100 years, the behaviour of enzymes had been explained by the "lock-and-key" mechanism developed by pioneering German chemist Emil Fischer. Fischer thought that the chemicals
REVIEW. Signal transduction is a fundamental biological process that is. Molecular signatures of G-protein-coupled receptors
 EVIEW doi:10.1038/nature11896 Molecular signatures of G-protein-coupled receptors A. J. Venkatakrishnan 1, Xavier Deupi 2, Guillaume Lebon 1,3,4,5, Christopher G. Tate 1, Gebhard F. Schertler 2,6 & M.
EVIEW doi:10.1038/nature11896 Molecular signatures of G-protein-coupled receptors A. J. Venkatakrishnan 1, Xavier Deupi 2, Guillaume Lebon 1,3,4,5, Christopher G. Tate 1, Gebhard F. Schertler 2,6 & M.
Reception The target cell s detection of a signal coming from outside the cell May Occur by: Direct connect Through signal molecules
 Why Do Cells Communicate? Regulation Cells need to control cellular processes In multicellular organism, cells signaling pathways coordinate the activities within individual cells that support the function
Why Do Cells Communicate? Regulation Cells need to control cellular processes In multicellular organism, cells signaling pathways coordinate the activities within individual cells that support the function
Gene regulation I Biochemistry 302. Bob Kelm February 25, 2005
 Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Gene regulation I Biochemistry 302 Bob Kelm February 25, 2005 Principles of gene regulation (cellular versus molecular level) Extracellular signals Chemical (e.g. hormones, growth factors) Environmental
Model Worksheet Student Handout
 Introduction Despite the complexity of life on Earth, the most important large molecules found in all living things (biomolecules) can be classified into only four main categories: carbohydrates, lipids,
Introduction Despite the complexity of life on Earth, the most important large molecules found in all living things (biomolecules) can be classified into only four main categories: carbohydrates, lipids,
BIBC 100. Structural Biochemistry
 BIBC 100 Structural Biochemistry http://classes.biology.ucsd.edu/bibc100.wi14 Papers- Dialogue with Scientists Questions: Why? How? What? So What? Dialogue Structure to explain function Knowledge Food
BIBC 100 Structural Biochemistry http://classes.biology.ucsd.edu/bibc100.wi14 Papers- Dialogue with Scientists Questions: Why? How? What? So What? Dialogue Structure to explain function Knowledge Food
d ^dzd K DZ/ '^ ^ '^ K '^ / '^ / DZ/ W / ^ d Eled ^ d Eldd dd d e t d d e de ^ '^ de ddl '^
 DZ/, ^ : ' t ' D ^ E > E / D, ^ : ' W > E/D, E/, dd Z d dde Dddeed ddee ddd ddd dede E /, D d ^dzd K DZ/ '^ ^ '^ K '^ / '^ / DZ/ W / ^ d Eled ^ d Eldd dd d e t d d e de ^ '^ de ddl '^ d d '^ t '^ d /EdZKhd/KE
DZ/, ^ : ' t ' D ^ E > E / D, ^ : ' W > E/D, E/, dd Z d dde Dddeed ddee ddd ddd dede E /, D d ^dzd K DZ/ '^ ^ '^ K '^ / '^ / DZ/ W / ^ d Eled ^ d Eldd dd d e t d d e de ^ '^ de ddl '^ d d '^ t '^ d /EdZKhd/KE
Supplementary Figure 1
 Supplementary Figure 1 a Tyr191 Trp265 4-5 Å Tyr268 ~14 Å 4.3 Å 16 17 19 20 4.4 Å 11 15 Lys296 N 8 10 12 14 7.3 Å 18 Tyr191 4-5 Å 18 3.8-4 Å 8 10 12 11 17 16 19 ~ Å ~4 Å Tyr268 5.5-6.3 Å 14 N Lys296 15
Supplementary Figure 1 a Tyr191 Trp265 4-5 Å Tyr268 ~14 Å 4.3 Å 16 17 19 20 4.4 Å 11 15 Lys296 N 8 10 12 14 7.3 Å 18 Tyr191 4-5 Å 18 3.8-4 Å 8 10 12 11 17 16 19 ~ Å ~4 Å Tyr268 5.5-6.3 Å 14 N Lys296 15
Sunhats for plants. How plants detect dangerous ultraviolet rays
 Sunhats for plants How plants detect dangerous ultraviolet rays Anyone who has ever suffered sunburn will know about the effects of too much ultraviolet (UV) radiation, in particular UV-B (from 280-315
Sunhats for plants How plants detect dangerous ultraviolet rays Anyone who has ever suffered sunburn will know about the effects of too much ultraviolet (UV) radiation, in particular UV-B (from 280-315
Conformational Analysis
 Conformational Analysis C01 3 C C 3 is the most stable by 0.9 kcal/mole C02 K eq = K 1-1 * K 2 = 0.45-1 * 0.048 = 0.11 C04 The intermediate in the reaction of 2 has an unfavorable syn-pentane interaction,
Conformational Analysis C01 3 C C 3 is the most stable by 0.9 kcal/mole C02 K eq = K 1-1 * K 2 = 0.45-1 * 0.048 = 0.11 C04 The intermediate in the reaction of 2 has an unfavorable syn-pentane interaction,
CD Basis Set of Spectra that is used is that derived from comparing the spectra of globular proteins whose secondary structures are known from X-ray
 CD Basis Set of Spectra that is used is that derived from comparing the spectra of globular proteins whose secondary structures are known from X-ray crystallography An example of the use of CD Modeling
CD Basis Set of Spectra that is used is that derived from comparing the spectra of globular proteins whose secondary structures are known from X-ray crystallography An example of the use of CD Modeling
2: CHEMICAL COMPOSITION OF THE BODY
 1 2: CHEMICAL COMPOSITION OF THE BODY Although most students of human physiology have had at least some chemistry, this chapter serves very well as a review and as a glossary of chemical terms. In particular,
1 2: CHEMICAL COMPOSITION OF THE BODY Although most students of human physiology have had at least some chemistry, this chapter serves very well as a review and as a glossary of chemical terms. In particular,
NGF - twenty years a-growing
 NGF - twenty years a-growing A molecule vital to brain growth It is twenty years since the structure of nerve growth factor (NGF) was determined [ref. 1]. This molecule is more than 'quite interesting'
NGF - twenty years a-growing A molecule vital to brain growth It is twenty years since the structure of nerve growth factor (NGF) was determined [ref. 1]. This molecule is more than 'quite interesting'
Lecture 11: Protein Folding & Stability
 Structure - Function Protein Folding: What we know Lecture 11: Protein Folding & Stability 1). Amino acid sequence dictates structure. 2). The native structure represents the lowest energy state for a
Structure - Function Protein Folding: What we know Lecture 11: Protein Folding & Stability 1). Amino acid sequence dictates structure. 2). The native structure represents the lowest energy state for a
Protein Folding & Stability. Lecture 11: Margaret A. Daugherty. Fall Protein Folding: What we know. Protein Folding
 Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2003 Structure - Function Protein Folding: What we know 1). Amino acid sequence dictates structure. 2). The native structure represents
Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2003 Structure - Function Protein Folding: What we know 1). Amino acid sequence dictates structure. 2). The native structure represents
Structural Analysis and Dynamics of Retinal Chromophore in Dark and Meta I States of Rhodopsin from 2 H NMR of Aligned Membranes
 doi:10.1016/j.jmb.2007.03.046 J. Mol. Biol. (2007) 372, 50 66 Structural Analysis and Dynamics of Retinal Chromophore in Dark and Meta I States of Rhodopsin from 2 H NMR of Aligned Membranes Andrey V.
doi:10.1016/j.jmb.2007.03.046 J. Mol. Biol. (2007) 372, 50 66 Structural Analysis and Dynamics of Retinal Chromophore in Dark and Meta I States of Rhodopsin from 2 H NMR of Aligned Membranes Andrey V.
A Ligand Channel through the G Protein Coupled Receptor Opsin
 A Ligand Channel through the G Protein Coupled Receptor Opsin Peter W. Hildebrand 1. *, Patrick Scheerer 1., Jung Hee Park 1, Hui-Woog Choe 2, Ronny Piechnick 1, Oliver P. Ernst 1, Klaus Peter Hofmann
A Ligand Channel through the G Protein Coupled Receptor Opsin Peter W. Hildebrand 1. *, Patrick Scheerer 1., Jung Hee Park 1, Hui-Woog Choe 2, Ronny Piechnick 1, Oliver P. Ernst 1, Klaus Peter Hofmann
Three-layer ONIOM studies of the dark state of Rhodopsin: the protonation state of Glu181
 Accepted Manuscript Three-layer ONIOM studies of the dark state of Rhodopsin: the protonation state of Glu181 Katherine. F. Hall, Thom Vreven, Michael J. Frisch, Michael J. Bearpark PII: S0022-2836(08)00987-X
Accepted Manuscript Three-layer ONIOM studies of the dark state of Rhodopsin: the protonation state of Glu181 Katherine. F. Hall, Thom Vreven, Michael J. Frisch, Michael J. Bearpark PII: S0022-2836(08)00987-X
Membrane Protein Channels
 Membrane Protein Channels Potassium ions queuing up in the potassium channel Pumps: 1000 s -1 Channels: 1000000 s -1 Pumps & Channels The lipid bilayer of biological membranes is intrinsically impermeable
Membrane Protein Channels Potassium ions queuing up in the potassium channel Pumps: 1000 s -1 Channels: 1000000 s -1 Pumps & Channels The lipid bilayer of biological membranes is intrinsically impermeable
A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility
 proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility") (P&S Ch 5; Fer Ch 2, 9; Palm Ch 10,11; Zub Ch 9) A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility B.
(P&S Ch 5; Fer Ch 2, 9; Palm Ch 10,11; Zub Ch 9) A. Reaction Mechanisms and Catalysis (1) proximity effect (2) acid-base catalysts (3) electrostatic (4) functional groups (5) structural flexibility B.
Signal Transduction. Dr. Chaidir, Apt
 Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
Signal Transduction Dr. Chaidir, Apt Background Complex unicellular organisms existed on Earth for approximately 2.5 billion years before the first multicellular organisms appeared.this long period for
SUPPLEMENTARY INFORMATION
 Table of Contents Page Supplementary Table 1. Diffraction data collection statistics 2 Supplementary Table 2. Crystallographic refinement statistics 3 Supplementary Fig. 1. casic1mfc packing in the R3
Table of Contents Page Supplementary Table 1. Diffraction data collection statistics 2 Supplementary Table 2. Crystallographic refinement statistics 3 Supplementary Fig. 1. casic1mfc packing in the R3
Proteorhodopsin phototrophy in the ocean
 Nature 411, 786-789 (14 June 2001) doi:10.1038/35081051; Received 3 January 2001; Accepted 26 March 2001 Proteorhodopsin phototrophy in the ocean Oded Béjà, Elena N. Spudich, John L. Spudich, Marion Leclerc
Nature 411, 786-789 (14 June 2001) doi:10.1038/35081051; Received 3 January 2001; Accepted 26 March 2001 Proteorhodopsin phototrophy in the ocean Oded Béjà, Elena N. Spudich, John L. Spudich, Marion Leclerc
Molecular Basis of K + Conduction and Selectivity
 The Structure of the Potassium Channel: Molecular Basis of K + Conduction and Selectivity -Doyle, DA, et al. The structure of the potassium channel: molecular basis of K + conduction and selectivity. Science
The Structure of the Potassium Channel: Molecular Basis of K + Conduction and Selectivity -Doyle, DA, et al. The structure of the potassium channel: molecular basis of K + conduction and selectivity. Science
Proteins are not rigid structures: Protein dynamics, conformational variability, and thermodynamic stability
 Proteins are not rigid structures: Protein dynamics, conformational variability, and thermodynamic stability Dr. Andrew Lee UNC School of Pharmacy (Div. Chemical Biology and Medicinal Chemistry) UNC Med
Proteins are not rigid structures: Protein dynamics, conformational variability, and thermodynamic stability Dr. Andrew Lee UNC School of Pharmacy (Div. Chemical Biology and Medicinal Chemistry) UNC Med
Supplementary Information. Structural basis for precursor protein-directed ribosomal peptide macrocyclization
 Supplementary Information Structural basis for precursor protein-directed ribosomal peptide macrocyclization Kunhua Li 1,3, Heather L. Condurso 1,3, Gengnan Li 1, Yousong Ding 2 and Steven D. Bruner 1*
Supplementary Information Structural basis for precursor protein-directed ribosomal peptide macrocyclization Kunhua Li 1,3, Heather L. Condurso 1,3, Gengnan Li 1, Yousong Ding 2 and Steven D. Bruner 1*
Available online at ~DIRI=CT SCIENCE. Structure of Bovine Rhodopsin in a Trigonal Crystal Form
 doi:l 0.1016/j.jmb.2004.08.090 J. MoL BioL (2004) 343, 1409-1438 jib Available online at www.sciencedirect.com SCIENCE ~DIRI=CT ELSEVIER Structure of Bovine Rhodopsin in a Trigonal Crystal Form Jade Li
doi:l 0.1016/j.jmb.2004.08.090 J. MoL BioL (2004) 343, 1409-1438 jib Available online at www.sciencedirect.com SCIENCE ~DIRI=CT ELSEVIER Structure of Bovine Rhodopsin in a Trigonal Crystal Form Jade Li
Table 1. Crystallographic data collection, phasing and refinement statistics. Native Hg soaked Mn soaked 1 Mn soaked 2
 Table 1. Crystallographic data collection, phasing and refinement statistics Native Hg soaked Mn soaked 1 Mn soaked 2 Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell
Table 1. Crystallographic data collection, phasing and refinement statistics Native Hg soaked Mn soaked 1 Mn soaked 2 Data collection Space group P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 P2 1 2 1 2 1 Cell
Protein Folding & Stability. Lecture 11: Margaret A. Daugherty. Fall How do we go from an unfolded polypeptide chain to a
 Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2004 How do we go from an unfolded polypeptide chain to a compact folded protein? (Folding of thioredoxin, F. Richards) Structure - Function
Lecture 11: Protein Folding & Stability Margaret A. Daugherty Fall 2004 How do we go from an unfolded polypeptide chain to a compact folded protein? (Folding of thioredoxin, F. Richards) Structure - Function
Hole s Human Anatomy and Physiology Eleventh Edition. Chapter 2
 Hole s Human Anatomy and Physiology Eleventh Edition Shier Butler Lewis Chapter 2 1 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. CHAPTER 2 CHEMICAL BASIS OF
Hole s Human Anatomy and Physiology Eleventh Edition Shier Butler Lewis Chapter 2 1 Copyright The McGraw-Hill Companies, Inc. Permission required for reproduction or display. CHAPTER 2 CHEMICAL BASIS OF
SUPPLEMENTARY INFORMATION
 Figure S1. Secondary structure of CAP (in the camp 2 -bound state) 10. α-helices are shown as cylinders and β- strands as arrows. Labeling of secondary structure is indicated. CDB, DBD and the hinge are
Figure S1. Secondary structure of CAP (in the camp 2 -bound state) 10. α-helices are shown as cylinders and β- strands as arrows. Labeling of secondary structure is indicated. CDB, DBD and the hinge are
Central Dogma. modifications genome transcriptome proteome
 entral Dogma DA ma protein post-translational modifications genome transcriptome proteome 83 ierarchy of Protein Structure 20 Amino Acids There are 20 n possible sequences for a protein of n residues!
entral Dogma DA ma protein post-translational modifications genome transcriptome proteome 83 ierarchy of Protein Structure 20 Amino Acids There are 20 n possible sequences for a protein of n residues!
NMR, X-ray Diffraction, Protein Structure, and RasMol
 NMR, X-ray Diffraction, Protein Structure, and RasMol Introduction So far we have been mostly concerned with the proteins themselves. The techniques (NMR or X-ray diffraction) used to determine a structure
NMR, X-ray Diffraction, Protein Structure, and RasMol Introduction So far we have been mostly concerned with the proteins themselves. The techniques (NMR or X-ray diffraction) used to determine a structure
Supporting Information
 Supporting Information Ottmann et al. 10.1073/pnas.0907587106 Fig. S1. Primary structure alignment of SBT3 with C5 peptidase from Streptococcus pyogenes. The Matchmaker tool in UCSF Chimera (http:// www.cgl.ucsf.edu/chimera)
Supporting Information Ottmann et al. 10.1073/pnas.0907587106 Fig. S1. Primary structure alignment of SBT3 with C5 peptidase from Streptococcus pyogenes. The Matchmaker tool in UCSF Chimera (http:// www.cgl.ucsf.edu/chimera)
SUPPLEMENTARY INFORMATION
 www.nature.com/nature 1 Figure S1 Sequence alignment. a Structure based alignment of the plgic of E. chrysanthemi (ELIC), the acetylcholine binding protein from the snail Lymnea stagnalis (AchBP, PDB code
www.nature.com/nature 1 Figure S1 Sequence alignment. a Structure based alignment of the plgic of E. chrysanthemi (ELIC), the acetylcholine binding protein from the snail Lymnea stagnalis (AchBP, PDB code
Secondary Structure. Bioch/BIMS 503 Lecture 2. Structure and Function of Proteins. Further Reading. Φ, Ψ angles alone determine protein structure
 Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
Bioch/BIMS 503 Lecture 2 Structure and Function of Proteins August 28, 2008 Robert Nakamoto rkn3c@virginia.edu 2-0279 Secondary Structure Φ Ψ angles determine protein structure Φ Ψ angles are restricted
SUPPLEMENTARY INFORMATION
 doi:1.138/nature1737 Supplementary Table 1 variant Description FSEC - 2B12 a FSEC - 6A1 a K d (leucine) c Leucine uptake e K (wild-type like) K (Y18F) K (TS) K (TSY) K288A mutant, lipid facing side chain
doi:1.138/nature1737 Supplementary Table 1 variant Description FSEC - 2B12 a FSEC - 6A1 a K d (leucine) c Leucine uptake e K (wild-type like) K (Y18F) K (TS) K (TSY) K288A mutant, lipid facing side chain
Rex-Family Repressor/NADH Complex
 Kasey Royer Michelle Lukosi Rex-Family Repressor/NADH Complex Part A The biological sensing protein that we selected is the Rex-family repressor/nadh complex. We chose this sensor because it is a calcium
Kasey Royer Michelle Lukosi Rex-Family Repressor/NADH Complex Part A The biological sensing protein that we selected is the Rex-family repressor/nadh complex. We chose this sensor because it is a calcium
SUPPLEMENTARY INFORMATION
 doi:10.1038/nature11054 Supplementary Fig. 1 Sequence alignment of Na v Rh with NaChBac, Na v Ab, and eukaryotic Na v and Ca v homologs. Secondary structural elements of Na v Rh are indicated above the
doi:10.1038/nature11054 Supplementary Fig. 1 Sequence alignment of Na v Rh with NaChBac, Na v Ab, and eukaryotic Na v and Ca v homologs. Secondary structural elements of Na v Rh are indicated above the
Subhodeep Moitra 1, Kalyan C Tirupula 2, Judith Klein-Seetharaman 2* and Christopher James Langmead 1*
 Moitra et al. BMC Biophysics 2012, 5:13 RESEARCH ARTICLE Open Access A minimal ligand binding pocket within a network of correlated mutations identified by multiple sequence and structural analysis of
Moitra et al. BMC Biophysics 2012, 5:13 RESEARCH ARTICLE Open Access A minimal ligand binding pocket within a network of correlated mutations identified by multiple sequence and structural analysis of
Lecture 2 and 3: Review of forces (ctd.) and elementary statistical mechanics. Contributions to protein stability
 and elementary statistical mechanics. Contributions to protein stability") Lecture 2 and 3: Review of forces (ctd.) and elementary statistical mechanics. Contributions to protein stability Part I. Review of forces Covalent bonds Non-covalent Interactions: Van der Waals Interactions
Lecture 2 and 3: Review of forces (ctd.) and elementary statistical mechanics. Contributions to protein stability Part I. Review of forces Covalent bonds Non-covalent Interactions: Van der Waals Interactions
Homology models of the tetramerization domain of six eukaryotic voltage-gated potassium channels Kv1.1-Kv1.6
 Homology models of the tetramerization domain of six eukaryotic voltage-gated potassium channels Kv1.1-Kv1.6 Hsuan-Liang Liu* and Chin-Wen Chen Department of Chemical Engineering and Graduate Institute
Homology models of the tetramerization domain of six eukaryotic voltage-gated potassium channels Kv1.1-Kv1.6 Hsuan-Liang Liu* and Chin-Wen Chen Department of Chemical Engineering and Graduate Institute
4 Examples of enzymes
 Catalysis 1 4 Examples of enzymes Adding water to a substrate: Serine proteases. Carbonic anhydrase. Restrictions Endonuclease. Transfer of a Phosphoryl group from ATP to a nucleotide. Nucleoside monophosphate
Catalysis 1 4 Examples of enzymes Adding water to a substrate: Serine proteases. Carbonic anhydrase. Restrictions Endonuclease. Transfer of a Phosphoryl group from ATP to a nucleotide. Nucleoside monophosphate
The Signaling Pathway of Rhodopsin
 Article The Yifei Kong 1 and Martin Karplus 1,2, * 1 Department of Chemistry and Chemical Biology, Harvard University, Cambridge, MA 02138, USA 2 Laboratoire de Chimie Biophysique, ISIS, Université Louis
Article The Yifei Kong 1 and Martin Karplus 1,2, * 1 Department of Chemistry and Chemical Biology, Harvard University, Cambridge, MA 02138, USA 2 Laboratoire de Chimie Biophysique, ISIS, Université Louis
Overview of the Accountancy Profession in China
 2016I1-024 Overview of the Accountancy Profession in China (Released by CICPA, Updated on January 21, 2016) /K D d // / W d ///^ d d d /s^ / ^ e s^ W D & e s/^ E ^ e s///d^ W e s///y W dd /y/ dd y/ dd
2016I1-024 Overview of the Accountancy Profession in China (Released by CICPA, Updated on January 21, 2016) /K D d // / W d ///^ d d d /s^ / ^ e s^ W D & e s/^ E ^ e s///d^ W e s///y W dd /y/ dd y/ dd
BA, BSc, and MSc Degree Examinations
 Examination Candidate Number: Desk Number: BA, BSc, and MSc Degree Examinations 2017-8 Department : BIOLOGY Title of Exam: Molecular Biology and Biochemistry Part I Time Allowed: 1 hour and 30 minutes
Examination Candidate Number: Desk Number: BA, BSc, and MSc Degree Examinations 2017-8 Department : BIOLOGY Title of Exam: Molecular Biology and Biochemistry Part I Time Allowed: 1 hour and 30 minutes